Киргизия, Талас


СДЕЛАЙТЕ СВОИ УРОКИ ЕЩЁ ЭФФЕКТИВНЕЕ, А ЖИЗНЬ СВОБОДНЕЕ
Благодаря готовым учебным материалам для работы в классе и дистанционно
Скидки до 50 % на комплекты
только до
Готовые ключевые этапы урока всегда будут у вас под рукой
Организационный момент
Проверка знаний
Объяснение материала
Закрепление изученного
Итоги урока

Была в сети 02.03.2022 09:49

Медетбекова Мээрим Болоткановна
Преподаватель кафедры "Естествознания"
27 лет
Местоположение
Лекции по геологии
Категория:
География
02.03.2022 09:48

Просмотр содержимого документа
«Лекции по геологии»
Лекция 15-16
Основные этапы развития жизни на Земле.
1. История развития Земли в докембрии.
2.Раннепалеозойский этап развития Земли.
3.Позднепалеозойский этап развития Земли.
4.История развития Земли в мезозое и кайнозое.
Наша планета образовалась около 4,5 млрд. лет назад. Поверхность Земли была в то время очень горячей, и никакой жизни на ней не существовало. Самым древним остаткам живых организмов, которые чем-то походили на современных бактерий, предположительно 3,5 млрд. лет. Заглянуть в прошлое Земли, проследить этапы развития жизни помогает палеонтология — наука о вымерших растениях и животных, чьи окаменевшие остатки сохранились в толщах осадочных пород.
Палеонтология развивалась рука об руку с геохронологией, изучающей геологическую историю Земли. Изучая строение, условия возникновения и напластования осадочных пород и научившись определять их абсолютный возраст, ученые создали геохронологическую шкалу, разделив геологическую историю Земли на ряд периодов. Продолжительность этих периодов измеряется миллионами лет, все они разделены на 5 эр: архей, протерозой, палеозой, мезозой, кайнозой. Архей — самая древняя эра, а в кайнозое мы живем.
Зарождение жизни
На заре своего существования Земля представляла собой раскаленный безжизненный шар, и только 3,5 млрд. лет назад она остыла настолько, что на ее поверхности образовалась твердая земная кора. Пары воды, содержавшиеся в первичной атмосфере, сконденсировались и стали выпадать в виде дождей, образовав древний океан, над которым возвышался единственный материк, впоследствии расколовшийся на части. В земной коре происходили постоянные изменения, сопровождавшиеся грандиозными климатическими колебаниями. Именно в океане появились первые живые организмы, напоминавшие бактерий, а потом и одноклеточные водоросли.

В древнейшей атмосфере Земли не было кислорода, однако водоросли были способны использовать энергию солнечного света для производства питательных веществ. При этом они выделяли кислород, который стал накапливаться в воздухе, создав условия для возникновения более сложно устроенных организмов.
Появление кислорода оказалось губительным для организмов, привыкших к бескислородной среде, так произошло первое на Земле вымирание. Прошло немало времени, прежде чем в океане появились первые многоклеточные организмы. Это были губки, медузы и организмы, напоминающие червей. Их остатки обнаружены в породах, возраст которых насчитывает около 600 млн. лет.
Развитие жизни на Земле было тесно связано с передвижением материков, которое влекло за собой изменения климата и уровня Мирового океана.
Раннепалеозойский (каледонский) этап развития Земли приходится на первую половину палеозойской эры. Он состоит из кембрийского, ордовикского и силурийского периодов общей продолжительностью 170±10 млн. лет. Широкое развитие животного и растительного мира, многие представители которых имели известковые или кремнистые раковинки и скелеты, позволяет произвести дробное деление этого отрезка геологической истории.
Геологические события раннего палеозоя. Развитие земной коры в раннем палеозое протекало различно на платформах и в геосинклиналях. Дифференциация тектонического режима, начавшаяся с позднего протерозоя, еще больше усилилась. Докембрийские платформы испытывали преимущественно медленные нисходящие вертикальные движения. Пониженные участки затопляются мелководными морями, где накапливались пески, глинистые и карбонатные илы. Иногда возникали лагунные условия и формировались осадочные породы химического происхождения: каменная соль, гипс, ангидрит.
Постепенно в прогибание втягивались все новые и новые территории платформы, образовались области площадью в несколько миллионов квадратных километров. Возникали плиты, мощность осадочного чехла в их пределах составляла 3 - 4 км. На Восточно-Европейской платформе оформилась Русская плита, на Северо-Американской - плита Великих равнин и плита Мидконтинент, на Сибирской платформе - Ангаро-Ленская плита и т. д. Отдельные участки платформ - щиты - не испытывали такого прогибания и оставались крупной островной сушей, это, например, Балтийский и Украинский щиты Восточно-Европейской платформы. Со щитов обломочный материал поступал в прилегающие моря.
В раннем палеозое платформы неоднократно испытывали наступание и отступание моря. Обычно считают, что "пики" морской трансгрессии приходятся на средний кембрий, средний ордовик и ранний силур. Между этими трансгрессиями проявлялись регрессии моря, и территория суши увеличивалась.
В геосинклиналях геологические процессы носили более напряженный характер. Западная и Восточная Сибирь, север Скандинавского полуострова, Средиземноморье, Кордильеры и Анды, Восточная Австралия - все это были геосинклинали, представляющие собой морские пространства с большим количеством островов. В кембрии и ордовике геосинклинали испытывали резко дифференцированные вертикальные движения многокилометровой амплитуды. Узкие вытянутые поднятия (геоантиклинали) соседствовали с такими же линейными прогибами, где накапливались галечники, пески, глины карбонатные илы, преобразованные в дальнейшем в конгломераты, песчаники, глинистые сланцы, известняки, доломиты. Большим распространением пользовались эффузивные породы, указывающие на интенсивную вулканическую деятельность. В геосинклинальных прогибах мощность накопленных осадков достигала 8 - 12 км, тогда как на смежных поднятиях она не превышала первые километры.
В силуре размеры геосинклинальных морей резко сократились. Глобальное сокращение площади морей и океанов объясняется тем, что в конце силура особенно интенсивно проявился диастрофизм каледонской тектономагматической эпохи. В результате многие геосинклинали преобразовались в платформы, которые в последующем уже не испытывали активных тектонических движений и вулканизма.
Образовавшиеся в конце раннего палеозоя устойчивые области земной коры получили название эпикаледонских (рис. 10). Они, как правило, располагались по перифериям докембрийских платформ, увеличивая тем самым их общую площадь. Однако во многих местах геосинклинальный режим сохранился - Средиземноморье, Западная и Восточная Сибирь, Кордильеры и Анды, Восточная Австралия продолжали свое геосинклинальное развитие и в позднем палеозое.

Рис. 10. Схема современного положения каледонид. 1 - эпикаледонские платформы: 1 - Северо-Грендландская, 2 - Грампианская, 3 - Иннуитская, 4 - Центрально-Казахстанская, 5 - Алтае-Саянская, 6 - Наньшанская, 7 - Катазиатская; 2 - области более ранней консолидации; 3 - блоки раздробленной древней Китайской платформы
Населенные океаны и безжизненная суша - такой застали бы мы нашу планету в раннем палеозое. Несмотря на многообразие жизни, она практически вся сосредоточилась в водных бассейнах. Поэтому, чтобы познакомиться с ней, придется опуститься на дно раннепалеозойского океана.
На илистом дне мы увидели бы странных существ, чем-то напоминающих мокриц, но гораздо больших размеров. Это - трилобиты, особый класс членистоногих животных, давно уже исчезнувшие с лика Земли. Тело трилобитов расчленялось на три сегмента (щита): головной, туловищный и хвостовой (рис. 11, а). Со стороны спины животное защищено тонким хитиновым панцирем (до 1 мм). В кембрии трилобиты обладали хорошо развитым головным щитом и слабо развитым хвостовым. Они составляли до 60% всей известной кембрийской фауны. Органы зрения у них были недоразвиты или же отсутствовали. В ордовике у трилобитов хорошо развиваются хвостовой щит и глаза. Животные приобрели способность свертываться, подтягивать задний щит к головному, защищая тем самым нежное брюшко от врагов. В силуре продолжается совершенствование органов зрения, у трилобитов появились большие сетчато-сложные глаза, которые позволяли ориентироваться по ультрафиолетовым и инфракрасным сигналам.

Рис. 11. Некоторые характерные представители раннепалеозойской фауны. Трилобиты: Илленус и Азафус (а); граптолит Диктионема (в); брахиоподы: Пентамерус бореалис (б) и Ортис каллиграмма (г)
Трилобиты вели исключительно придонный образ жизни. Ползая, они захватывали ртом, который находился на нижней части головного щита, ил с пищевыми частичками. Животные достигали в длину 3 - 10 см, известны гиганты длиной до 70 см. Расцвет трилобитов приходится на кембрийский и ордовикский периоды. В конце силура количество родов трилобитов резко сокращается. В этом были повинны другие обитатели ранне-палеозойских морей и океанов - головоногие моллюски (наутилоидеи), для которых трилобиты были излюбленным блюдом: своими щупальцами с присосками они хватали трилобитов, притягивали к роговым челюстям клюва и перемалывали на терке из многочисленных рядов пластин и крючков. В то время пищи для хищных моллюсков было много, и они стали увеличиваться в своих размерах; известны раковины головоногих до 2 и даже 4,5 м длины. Урон, нанесенный трилобитам, оказался роковым: в позднем палеозое сохранились единичные представители этой группы животных, а к концу палеозойской эры они полностью вымирают. Наутилоидеи же дожили до наших дней.
В силуре развиваются крупные ракообразные - отряд эвриптерид (ракоскорпионы). Эти животные имели удлиненное тело, достигавшее 1 и даже 3 м в длину.
В большинстве своем ракоскорпионы были хищниками. Не имея врагов, эвриптериды широко расселились не только в солоноватых, но и в пресных водоемах.
Наряду с трилобитами, головоногими моллюсками и ракоскорпионами океаны раннего палеозоя были заселены брахиоподами - животными, мягкое тело которых заключено в двустворчатую известковую раковину (рис. 11, б, г). С помощью хрящевидной ножки, выходившей между створками раковины, брахиоподы прикреплялись к морскому дну и ползали по нему. В кембрийском периоде эти животные составляли до 30% фауны. В океанах и морях раннего палеозоя встречались также губки, примитивные иглокожие (текоидеи и бластоидеи), кишечнополостные, черви, археоциаты.
Важным моментом в развитии органического мира было появление в раннем палеозое первых позвоночных животных. Наиболее примитивную группу полухордовых образовывали граптолиты - животные, практически вымершие к концу силура. Тонкое и длинное тело граптолита было заключено в хитиновую пленку, животные вели пассивный (планктонный) образ жизни, безвольно плавая в виде кистеобразной колонии в морских водах, лишь некоторые представители граптолитов приспособились к жизни на дне моря (рис. 11, в).
В прибрежно-морских условиях начинают развиваться первые бесчелюстные рыбообразные. В силуре уже появляются настоящие рыбы (акантоды). Тело их было покрыто чешуей, имелся внутренний хрящевой скелет.
Среди растений продолжали господствовать синезеленые и багряные водоросли, бактерии и грибы. В конце силура растения начинают робко выходить на сушу, появляясь в болотистых, часто заливаемых морем низинах. Эти растения образуют тип псилофитовидных. От горизонтального подземного стебля (корневища), который часто скрывался под дном водоема, вверх отходили вертикальные или стелющиеся побеги. Каждый' побег ветвился дважды (дихотомически). У одних псилофитовидных побеги были голыми, у других имелись уже зачаточные листики в виде шипов.
Таким образом, ранний палеозой был временем, когда органическая жизнь начинает бурно развиваться по сравнению с предыдущими этапами криптозоя (см. рис. 6). Этот "взрыв" жизни на Земле произошел сравнительно недавно - 500 - 600 млн. лет назад.
Несмотря на все многообразие органической жизни, Материки раннего палеозоя все еще представляли собой безжизненные пространства; кое-где могли виднеться лиловатого цвета колонии бактерий и грибов, да в прибрежных болотистых равнинах редели тонкие стволики псилофилитовых растений. Возможно, что выход живых организмов на сушу сдерживался недостатком кислорода в атмосфере, масса которого составляла всего около одной трети от его современного количества.
Климат раннего палеозоя характеризовался, вероятно, преобладанием теплых условий. В некоторых районах земного шара (Сибирь, Индия, Аравия) он был засушливый (аридный), о чем свидетельствуют соленосные отложения этих районов.
Позднепалеозойская история
Позднепалеозойский (герцинский) этап развития Земли включает три периода: девонский, каменноугольный и пермский - общей продолжительностью 170±10 млн. лет. Так же, как и ранний палеозой, поздний делится не только на периоды и эпохи, но и на века, отложения которых установлены практически на всех континентах.
Платформы растут. В геологическом отношении события позднепалеозойской истории сыграли важную роль в развитии Земли. Это выразилось в новом, очень значительном увеличении территории платформ.
В начале девона (400 млн. лет назад) докембрийские платформы и каледонские области испытывают, как правило, регрессию моря, которая началась еще в позднем силуре. В дальнейшем в среднем, позднем девоне и карбоне море неоднократно трансгрессирует на платформы и вновь отступает. Обычно платформенные морские бассейны носили мелководный характер. В них осаждались пески, глины, карбонатные илы. Последние часто целиком состояли из раковин беспозвоночных организмов, населявших позднепалеозойские океаны. В прибрежных районах платформ формируются угленосные осадки.
Докембрийские платформы испытывают сложные и разнонаправленные вертикальные движения, усилившие контраст между щитами и плитами, антеклизами и синеклизами. Территория плит, как правило, увеличивается. Так, например, значительно возросли размеры Русской плиты за счет вовлечения в прогибание соседних областей Балтийского и Украинского щитов. Растет и общая мощность осадочного чехла.
Каледонские области в начале позднего палеозоя (девонский период) обладали, в отличие от докембрийских платформ, четко выраженным горным рельефом. В межгорных впадинах накапливались мощные толщи красноцветных конгломератов с прослоями вулканических пород. Постепенно рельеф сглаживается, и в позднем девоне и в карбоне уже отлагались нормальные морские осадки.
В геосинклиналях в начале девона формировались грубообломочные породы (конгломераты), которые позднее сменялись песчано-глинистыми и карбонатными отложениями. В каменноугольном периоде сохранились условия для накопления чередующихся песчано-глинистых и карбонатных пород. Среди них в большом количестве встречаются прослои каменного угля. Во многих местах геосинклиналей в девоне и в карбоне интенсивно проявлялся вулканизм. В некоторых районах, например на Урале, метаморфизованные вулканические и кремнистые осадочные породы преобразовались в яшмы, слюдистые сланцы. Мощность отложений, накопленных в геосинклиналях, достигает 15 - 20 км.
В середине каменноугольного периода земная кора начинает испытывать новую волну складкообразовательных движений - герцинский тектогенез. Это была очень важная тектоно-магматическая эпоха в геологической истории Земли, проявившаяся на огромных территориях. На месте многих геосинклиналей возникают горы, поднятие испытывают и платформы. В пермский период всеобщее воздымание усиливается, и море почти повсеместно регрессирует. Пермский период - это время широкого развития суши: низменной на платформах и гористой в геосинклиналях. В связи с этим в Перми чаще всего накапливаются обломочные отложения с прослоями эффузивных пород в геосинклиналях. Важной особенностью пермского времени явилось формирование во многих местах земного шара каменной соли, причем мощность этих осадков достигает иногда 3 - 4 км. Соленосные образования пермского возраста известны на Восточно-Европейской платформе (Прикаспийская синеклиза), на севере Европы (Северо-Германская впадина), в Северной Америке (свита Биг-Блю).
Многие геосинклинали в результате позднепалеозойских фаз диастрофизма прекращают свое развитие и трансформируются в эпигерцинские платформы. Общая территория платформ при этом резко возрастает (рис. 12). Впервые в геологической истории Земли между возникшими герцинскими горными сооружениями и уже существовавшими докембрийскими платформами образуются узкие лентообразные прогибы, получившие название краевых, или передовых, прогибов. Наиболее крупные из них возникли на границе Урала и Восточно-Европейской платформы и на границе Аппалачей и Североамериканской платформы. Появление передовых прогибов знаменует качественно новые особенности геологической истории Земли.

Рис. 12. Схема современного положения герцинид. 1 - эпигерцинские платформы: 1 - Аппалачская, 2 - Южно-Американская или Патагонская, 3 - Западно-Европейская, 4 - Урало-Тяньшанская, 5 - Монголо-Охотская, 6 - Северо-Африканская или Антиатласская, 7 - Южно-Африканская или Капская, 8 - Восточно-Австралийская; 2 - области более ранней консолидации; 3 - передовые прогибы: А - Предаппалачский, Б - Предуральской
Образование эпигерцинских платформ значительно сократило территорию геосинклиналей. Герциниды в некоторых случаях явились своеобразным мостом между докембрийскими платформами. По мнению Б. Хизена и С. Кэри, в позднем палеозое платформенные массивы, образовавшие континенты, занимали до 80% поверхности суши Земли. Геосинклинальный режим сохранился в Средиземноморье, на Востоке Сибири, на западе Северной и Южной Америки.
Обновление животного мира. Рубеж между ранним и поздним палеозоем явился вехой в эволюции животных и растений. Одни представители животных вымирают, другие резко сокращают свое жизненное пространство и численность, на смену им появляются совершенно новые представители органического мира. Так, значительно сокращается (а в дальнейшем полностью вымирает) фауна трилобитов. Если в силуре насчитывалось 80 родов с 1200 видами, то в девоне сохранились лишь 11 родов с 105 видами. Еще более жесткая участь постигла граптолитов; из 32 силурийских родов в девоне сохранился лишь один род. Резко сократилось количество некоторых представителей иглокожих, наутилоидей, кораллов.
Из беспозвоночных наибольшего расцвета в позднем палеозое достигли брахиоподы, количество родов которых исчислялось многими сотнями (рис. 13, в, г). Развивались спирально закрученные головоногие моллюски (аммониты), простейшие, древние морские ежи, морские лилии, четырехлучевые и трубчатые кораллы. Широкое развитие в морях и океанах получают рыбы, в прибрежных лагунах - ракоскорпионы. Во второй половине позднего палеозоя на сушу выходят первые животные - земноводные, а в дальнейшем континенты завоевывают представители класса пресмыкающихся (рептилии).

Рис. 13. Некоторые характерные представители позднепалеозойской фауны. Панцирная рыба девона Птерихтус (а); пермская амфибия Какопс (б), брахиоподы: Циртоспирифер дизъюнктус (в), Пентамерус башкирикус (г)
В растительном мире также произошли серьезные изменения. Если в океанах и морях господство сохраняется за водорослями (синезелеными, багряными, харовыми), то на суше главенствующую роль начинают играть высшие растения: псилофиты, плауны, хвощи, папоротники. Поздний палеозой явился временем активного наступления на материки не только животных, но и растений. Однако процесс завоевания суши происходил постепенно.
Обновление животного мира, по мнению ученых, явилось следствием геологического преобразования Земли. Активные тектонические движения каледонской тектоно-магматической эпохи в конце раннего палеозоя вызвали образование во многих местах земного шара горных систем, резко сократились площади океанических бассейнов, изменилась циркуляция воздушных масс, климат стал засушливее, континентальнее. Столь существенное изменение ландшафта суши и перераспределение морских бассейнов планеты, по-видимому, оказало решающее влияние и на развитие животных и растений. Таким образом, смена условий существования на рубеже раннего и позднего палеозоя повлекла за собой и смену фауны и флоры.
Девон - век рыб. Ландшафт девонского времени мало чем отличался от силурийского пейзажа. В начале девонского периода континенты были заселены псилофитами лишь в прибрежных заболоченных районах, животные полностью отсутствовали. Основные события органической жизни происходили в океанах, где господствовали рыбы.
Девон называют иногда веком рыб. Вначале это были так называемые пластинокожие, или "панцирные", рыбы. Своеобразный панцирь покрывал голову и переднюю часть туловища (рис. 13, а). Панцирные рыбы были хищниками, они имели челюсть с зазубренными костными пластинками. Объектом их питания были в основном ракоскорпионы. Превосходя их в маневренности и в силе, панцирные рыбы окончательно оттеснили ракоскорпионов в лагуны и в прибрежные пресные водоемы. В середине девона появляются первые костные рыбы, разделившиеся в процессе эволюции на три ветви: лучеперые, кистеперые и двоякодышащие. В девоне преобладали кистеперые - хищные рыбы с сильными зубами. От лучеперых произошли современные акулы и скаты. Двоякодышащие рыбы известны своей способностью жить в засушливых условиях.
Долгое время считалось, что все представители древних девонских рыб вымерли и сохранились до наших дней лишь в ископаемом виде. В 1952 г. у побережья Южной Африки и у берегов Мадагаскара был обнаружен живой представитель кистеперых рыб - целакант, почти не отличавшийся от своих древних предков. У целаканта были сильные плавники, которые выполняли функцию ног. Тем самым было подтверждено, что именно кистеперые дали начало амфибиям - первым позвоночным животным, приспособившимся к жизни на суше.
В позднем девоне от кистеперых рыб произошли стегоцефалы ("крышеголовые"), относимые к земноводным животным - амфибиям (рис. 13, б). Это были первые четвероногие обитатели суши. Во многом жизнь стегоцефалов была связана с водой. Они размножались и выводили личинки в воде, подобно своим дальним потомкам - современным лягушкам и саламандрам. Стегоцефалы имели сплошную костную крышу черепа, вели хищный образ жизни, питались в основном рыбой.
В конце девона на суше появляются первые пауки и клещи. Намечается, таким образом, своеобразная "трансгрессия" животных на материк.
Во второй половине девона от псилофитовидных растений происходят плауновидные, папортниковидные. В позднем девоне псилофиты почти все исчезают и континенты захватывает археоптерисовая флора, названная так по преобладанию в ней папоротника археоптериса. Кроме плаунов и папоротников, в состав этой флоры входили также и первые семенные растения. Флора девонского времени приспосабливается к воздушной среде, к увеличению в воздухе содержания кислорода. У растений появляется кожица с устьицами - приспособление, предохраняющее их от высыхания. Тело растений дифференцируется на стебель, корни и листья, выполняющие различные функции.
Дремучие леса карбона. В каменноугольном периоде создались чрезвычайно благоприятные условия для развития наземной растительности. Теплый влажный климат господствовал на значительных пространствах земного шара. В атмосфере резко увеличилось содержание кислорода. Изобилие осадков приводило к заболачиванию громадных территорий, которые покрывались дремучими лесами, состоящими из гигантских древовидных - плауновых, хвощей и папоротников (лепидодендроны, сигиллярии, каламиты). Стволы деревьев до 30 - 40 м в высоту и до 2 м в поперечнике обвивали вьющиеся лианообразные растения с гибкими стеблями и колючками-крючками, цеплявшимися за стволы и сучья. В лесах карбона появились и первые голосеменные растения. По своей непроходимости каменноугольные леса были подобны современным мангровым зарослям болотистых тропических побережий. Густое сплетение сучьев и стволов деревьев создавало сумеречные дебри.
Душная, насыщенная тяжелыми запахами атмосфера царила в каменноугольных лесах. Травянистая растительность с мясистыми листьями и яркими крупными цветами, отмирая, формировала мощные залежи торфа. Под давлением вышележащих пород и повышенной температуры они преобразовывались сначала в бурые, а затем в каменные угли и антрациты. В условиях болот и озер чаще возникали бурые угли (лимническое угленакопление), тогда как в прибрежно-морских районах обычно формировались каменные угли и антрациты (паралическое угленакопление).
Резкое увеличение растительной биомассы на континентах привело к усиленному потреблению углекислоты из атмосферы. Частично она превращалась в углерод, накапливавшийся в органическом веществе, частично в свободный кислород, уходивший в атмосферу. Увеличение содержания в атмосфере кислорода способствовало усилению процессов химического выветривания, в частности, окислению различных минералов.
Широкое развитие теплолюбивой и влаголюбивой флоры характерно для раннекаменноугольной эпохи, та флора получила название антракофитовой. В среднем и позднем карбоне антракофитовая флора дифференцируется на вестфальскую, тунгусскую и гондванскую. Первая сохранила особенности тропической флоры раннего карбона. Тунгусская флора присуща умеренным климатическим зонам и представлена кордаитовыми растениями и папоротниками. Гондванская флора еще более бедная по разнообразию видов, чем тунгусская. Для нее характерны травянистые хвощевидные, кордантовые и глоссоптериевые растения, произрастающие в более холодном климате, чем представители тунгусской флоры.
Леса карбона были заселены земноводными - стегоцефалами и саламандрообразными животными. Они вели малоподвижный образ жизни, часами лежали на дне болот и озер, высунув из воды лишь кончик носа, выжидая добычу. В каменноугольный период появляются первые пресмыкающиеся - котилозавры, сохранившие сплошную крышку черепа, как и земноводные. Пресмыкающиеся (рептилии) уже имели две важные особенности: роговой покров, предохраняющий тело от высыхания, и способность откладывать на суше яйца с плотной оболочкой и большим количеством желтка. Таким образом, связь с водой у позвоночных становится еще более слабой, они лучше приспосабливаются к наземному образу жизни.
Господство по количеству видов на материках в карбоне захватывают насекомые - многочисленные паукообразные, скорпионообразные, стрекозы, таракановые к др. Начиная с этого времени насекомые, относящиеся к типу членистоногих (к тому же типу, что и трилобиты), прочно доминируют на Земле как по количеству видов, так и по общему количеству особей. На долю членистоногих приходится 86% от общего количества всех видов животных, а на долю только насекомых 80%! В настоящее время на одного человека приходятся 250 млн. насекомых. Поистине Земля - планета насекомых. В некоторых же случаях эти мелкие представители членистоногих образуют огромные стаи, как, например, саранча. Ученые подсчитали, что отдельные стаи саранчи весят столько же, сколько могут весить медь, свинец и цинк, добытые человеком за 100 последних лет. Однако никогда в истории Земли насекомые не достигали таких гигантских размеров, как в карбоне. Стрекозы имели размах крыльев до 100 см, тараканы достигали 50 см в длину.
В морях и океанах каменноугольного периода происходят небольшие изменения по сравнению с девонским временем. Преобладают брахиоподы (в особенности представители отрядов спирифериды и продуктиды), головоногие моллюски, иглокожие (морские лилии и морские ежи). Среди рыб сокращается количество акантод и увеличивается число лучеперых.
Необычайно интенсивно развиваются морские простейшие организмы. Размеры некоторых из них, например фузулинид, достигают до 1 - 2 см, тогда как обычно они измеряются долями миллиметра. Фузулиниды, так же, как и другие представители отряда фораминифер, принимают существенное участие в образовании органогенных известняков каменноугольного возраста. В этих известняках впоследствии накапливались залежи нефти и газа. Представители простейших и брахиопод являются руководящей фауной для карбона.
Ледники в Африке. В позднем палеозое наряду с областями теплого тропического климата существовали и полярные зоны. Широкое развитие материковых ледников в позднем палеозое (средний, поздний карбон) достоверно установлено на юге Африки, в Австралии, Индии, Южной Америке, Антарктиде. Причем по характерным штрихам, бороздам, царапинам, которые оставляют ледники на горных породах при движении, по особенностям расположения моренных отложений можно восстановить направление движения позднепалеозойских ледников. Считают, что существовало несколько центров оледенения, где зарождались ледники и откуда они начинали движение.
В некоторых случаях создается впечатление, будто центры оледенения находились вне современных материков. Так, в Южной Африке, в районе г. Дурбан, штриховка на поверхности горных пород указывает на то, что льды двигались со стороны современного Индийского океана. Это говорит в пользу существования единого суперконтинента Гондваны в позднем палеозое. В этом случае центр оледенения мог быть расположен в прилегавшей непосредственно к югу Африки Антарктиде, Правда, имеются иные суждения. Французский геолог П. Фурмарье считает, например, что гипотетический центр оледенения, откуда надвигались ледники на район г. Дурбан, отделен в настоящее время от Африки глубинным разломом и скрыт под водами Индийского океана.
Оледенение позднего палеозоя было грандиозным. Судя по мощности накопленных морен (до 300 - 600 м), можно предположить, что центральные области Гондваны в карбоне были покрыты ледяным панцирем, толщина которого могла достигать 5 - 6 км. Периодически ледники частично таяли. В толщах хаотических моренных отложений появляются в такие периоды прослои ленточных глин. Причины позднепалеозойского оледенения многие ученые видят в ином расположении полюсов (южный полюс, например, располагался на юге Африки), в иной циркуляции воздушных масс и в относительно высоком гипсометрическом положении Гондваны над уровнем моря (известно, что чем выше мы поднимаемся, тем становится холоднее; в среднем на 1000 м подъема температура понижается на 3 - 5°C).
На рис. 14 изображена реконструированная палеогеографическая схема земного шара каменноугольного периода (около 300 млн. лет назад). Косой штриховкой выделены области тропического климата, для которых характерны антракофитовая и вестфальская флоры. В основном тропики тогда проходили вдоль северных и северо-восточных берегов субширотного океана Тетис, охватывая современные районы Европы, частично Средней Азии, запад Северной Америки, севера Африки, севера и запада Южной Америки, предполагаемое положение экватора в то время показано на рисунке тонкой сплошной линией. Точками на рисунке выделены области оледенения. Они располагались в центре Гондваны. Стрелки указывают направление движения ледников. Площадь позднепалеозойского оледенения была необычайно велика. Это породило сомнение в возможности существования таких грандиозных ледников. Некоторые ученые считают даже, что на Земле не хватило бы воды для формирования столь огромных ледниковых масс. Несомненно одно: в позднем палеозое существовала четко выраженная климатическая зональность. Области полярного климата сменялись умеренным климатическим поясом, который в свою очередь переходил в тропический пояс.

Рис. 14. Палеогеографическая схема каменноугольного периода. По Д. Тэрлингу и М Тарлинг (1973)
Пермские рептилии. Значительное изменение климата на Земле произошло в пермский период. Активные тектонические движения герцинской эпохи диастрофизма, начавшие проявляться уже в среднем карбоне, привели в начале позднего карбона к завершению геосинклинального режима развития во многих районах земного шара. На месте геосинклиналей вырастают горные хребты. Урал, Западная Сибирь, Средняя Азия, Западная Европа, восток Северной Америки, восток Австралии - все это были молодые горные страны, возникшие в каменноугольном периоде. Моря, затоплявшие прежде эти территории, регрессируют, освобождая огромные пространства, или мелеют, превращаясь в гигантские лагуны. Характерной чертой пермского времени является накопление континентальных обломочных пород, имеющих красный цвет, и хемогенных лагунных отложений (каменная соль, гипс, ангидрит, доломит).
Сокращение морских бассейнов и резкое увеличение гористой суши повлияло на климат в сторону усиления его континентальности. Пышные леса карбона, не получая достаточной влаги, постепенно вырождаются, уступая место жарким пустыням. В связи с этим в поздней перми резко меняется флора: сокращается количество влаголюбивых хвощей, папоротников, плаунов, шире распространяются голосеменные растения - хвойные, цикадовые и гингковые, которые приспособились к существованию в засушливых условиях.
Значительные изменения происходят и в животном царстве. На суше земноводные постепенно отступают под напором рептилий. Амфибии продолжают развиваться в тропических областях (некоторые районы Северной Америки, ю. Европы, Юго-Восточная Азия). Многие амфибии приспосабливаются к жизни в дельтах рек, в лагунах и даже в более сухих местах. Рептилии, которые не нуждались в воде при размножении, захватывают в первую очередь пустынные, полупустынные и степные пространства.
Развитие наземной фауны позвоночных шло, по-видимому, в нескольких сравнительно обособленных центрах. Одним из таких центров считают Северную Америку (Техас), откуда известны богатейшие находки ранне-пермских амфибий (стегоцефалов) и рептилий. Последние были более разнообразны. Кроме типичных рептилий, среди них обнаружены примитивные пресмыкающиеся (котилозавры), имеющие некоторые сходные черты со стегоцефалами. Особенности захоронения этих животных указывают на то, что пермская фауна Техаса обитала в основном по берегам небольших озер, болот, рек, где обильная травянистая растительность давала пищу крупным травоядным рептилиям.
Другим возможным центром развития пермских позвоночных считают Южную Африку. Здесь обнаружены остатки котилозавров, териодонтов и дицинодонтов. Котилозавры - травоядные массивные животные, достигавшие 3 м в длину; их кожа была покрыта характерными роговыми выростами, создававшими своеобразный защитный панцирь.
Распространение рептилий из центров возникновения происходило по двум основным путям: из Северной Америки через Западную Европу до современного Урала, а из Южной Африки на северо-восток через Индию и Среднюю Азию в Восточную Европу и Великобританию и на юго-запад - в Южную Америку. В пределах СССР аналогичная фауна обнаружена в районах р. Северной Двины (комплекс фауны сходный с североамериканской) и в районах г. Тетюши и г. Очеры (комплекс фауны, тождественной южноафриканской).
В морях и океанах пермского времени также происходят существенные изменения. Если в течение всего пермского периода продолжают развиваться фузулиниды, губки, головоногие моллюски, кораллы, то конец перми для многих из них явился роковым рубежом. Выбирают фузулиниды, некоторые кораллы (табуляты, четырехлучевые кораллы), некоторые головоногие моллюски из наутилоидей и аммоноидей, некоторые иглокожие. Полностью вымирают также акантоды, древние лучеперые рыбы, палеозойские группы кистеперых и двоякодышащих рыб. Резко сокращается количество хрящевых рыб. Таким образом, конец позднего палеозоя, ознаменовавшийся образованием новых эпигерцинских платформ, отличается также и значительным обновлением флоры и фауны, что опять-таки явилось прямым следствием изменения геологической и географической обстановок в конце палеозоя.
Мезозойская история
Мезозойский этап развития Земли богат интересными геологическими событиями: раскалываются суперконтиненты Лавразия и Гондвана, отдельные блоки литосферы, соответствующие современным материкам, постепенно отдаляются друг от друга, давая начало новым океаническим впадинам. Принципиальные изменения происходят и в животном мире: появляются первые млекопитающие и птицы, в конце мезозоя вымирают динозавры. Все эти изменения происходят на протяжении мезозойской эры (170±10 млн. лет), состоящей из трех периодов: триасового, юрского и мелового.
Земной шар "трещит по швам". Начало мезозойского этапа знаменуется широкой регрессией морей и океанов. Рост размеров суши, начавшийся еще в пермском периоде за счет горообразовательных процессов, продолжался и в триасе. На обширных территориях платформ преобладал континентальный (геократический) режим, поэтому накапливались в основном обломочные красно-цветные отложения (пески, глины). В Азии имеются сероцветные обломочные осадки триаса с включением прослоев каменного угля.
Триасовый период - начало раскола Гондваны. В различных ее районах возникают глубинные разломы. Первоначальное положение их предопределило конфигурацию будущих материков южного полушария. По-видимому, ослабленные зоны, которые явились местом заложения разломов, стали намечаться в едином теле Гондваны еще в позднем палеозое. Это был общепланетарный процесс. Крупные разломы рассекли в то время и древнюю Сибирскую платформу. В перми вдоль них происходило излияние базальтовых лав, которое активно продолжалось и в триасе. Потоки базальта толщиной до 2,5 км покрыли территорию площадью свыше 500000 км2, образовав траппы. В триасовом периоде начинают проявлять активность и некоторые другие глубинные разломы.
С конца позднего палеозоя намечаются ослабленные зоны, обозначившие границы Африки, Индии, Австралии и Антарктиды. Эти зоны проявились образованием сравнительно узких, вытянутых прогибов, которые заполнялись главным образом обломочным материалом. В триасе прогибание на западной и южной окраинах Африканского континента продолжалось, формировались континентальные песчаные отложения. Между Африкой и Мадагаскаром существовал узкий морской залив, отражая стремление моря заполнить пониженный рельеф ослабленных зон Гондваны.
В конце триаса происходит интенсивная вспышка вулканизма. Глубинные разломы, "притаившиеся" в недрах Земли и постепенно "набиравшие силы", проявились в полной мере. По трещинам с глубины в несколько сот километров на поверхность устремились базальтовые лавы. Заполняя понижения в рельефе, они накапливались, образуя многосотметровые толщи. Под их давлением прогибание усиливается, из недр выдавливаются новые порции лав, которые, излившись, увеличивали давление на глубинные сферы Земли. К середине юрского периода вспышка вулканизма угасает. За это время образовалась лавовая толща, достигавшая в ряде районов (впадина Замбези, флексура Лебомбо) мощности 8 км. Одновозрастные лавы встречаются в Индии (траппы Радж-Махал), в Австралии (долериты о. Тасмании), на Среднем Востоке (траппы Йемена), в Антарктиде (Феррарские долериты).
Образование и активное проявление глубинных разломов ослабляет связь между отдельными блоками литосферы Гондваны. Намечается отделение Африки, Индии, Австралии и Антарктиды, между ними закладывается морской бассейн, превратившийся в дальнейшем в Индийский океан. Таким образом, возраст заложения последнего можно ориентировочно оценить в 160 млн. лет. Связь Африки с Южной Америкой в триасе была еще устойчивой.
Юрский период характеризуется постепенным развитием трансгрессии моря, максимум которой приходится на первую половину поздней юры (келловей, Оксфорд). Море захватывает запад Северной Америки, почти всю; Европу, Западную и Восточную Сибирь. В пределах Гондваны наступание моря идет со стороны формирующегося Индийского океана.
В конце юры вновь усиливаются движения по глубинным разломам, что сопровождается новым излиянием базальтовых лав, которые известны в Ливане, Сирии, Мозамбике. Начинают проявляться глубинные разломы на границе Африки и Южной Америки, в пределах последней накапливается толща лав мощностью до 1 км (лавы Сьерра-Гераль). В юрское время раскрывается впадина Северной Атлантики:
Активные тектонические движения конца юрского периода привели к некоторой регрессии моря, обмелению бассейнов и возникновению в ряде мест солеродных лагун. Новая трансгрессия достигает максимума в позднем мелу. Море покрывает значительную часть Лавразии и Гондваны. В мелу происходит третья вспышка магматической активности, сопровождавшей раскол Гондваны. Мощные лавовые потоки до 2 - 3 км образовывались в Африке, в Индии (Деканские траппы). К концу мела Индийский океан был уже четко выражен, хотя и не достиг современных размеров. Узким коленообразным морским проливом наметился Атлантический океан.
Мезозойский этап явился, таким образом, очень важным моментом в геологической истории Земли: с этого времени началось формирование современных континентов и океанов. Земной шар в буквальном смысле слова трещал по швам. Мощные внутренние силы Земли разрывали тонкую пленку земной коры, раздвигали ее "обрывки" в разные стороны, стремясь вырваться наружу в виде мощных лавовых излияний вдоль глубинных разломов.
Материки меняют свои очертания. Геологические события мезозойской эры привели к существенным изменениям в очертании материков. Единые супергигантские континенты - Лавразия и Гондвана распадаются (рис. 15).

Рис. 15. 'Расползание' материков в мезозое. По Д. Тарлингу и М. Тарлинг (1973). а - Лавразия и Гондвана в конце палеозойской эры; б - начало раскола суперконтинентов и их положение в конце триаса; в - расползающиеся континенты мела: 'раскрылся' Индийский океан и возникла узкая коленообразная лента Атлантики, 1 - протяженные геосинклинальные подвижные пояса; 2 - мелководные моря на континентах; 3 - глубинные разломы, определившие очертания современных материков
В позднем палеозое намечается ослабленная зона на западной границе современной Африки; в триасе глубинные разломы активизируются, вызывая проседание узких линейных зон земной коры и излияние базальтовых лав. В образовавшихся узких грабенах-рифтах первоначальная гранитная кора разрушается и перерабатывается, края грабенов под действием внутренних сил Земли удаляются друг от друга. Грабены-рифты заполняются морскими водами, возникают узкие внутриконтинентальные моря, напоминающие современные Красное море, Суэцкий залив, Калифорнийский залив, Аденский пролив. Подобные моря примерно 160 - 150 млн. лет назад распространяются к югу от океана Тетис, вклиниваясь между Индией и п-вом Сомали Африки.
Некоторое время спустя внутриконтинентальные моря возникают между Индией и Австралией. Несколько позже (120 млн. лет назад) морские воды образуют узкий пролив между Южной Америкой и Африкой. По данным английского ученого Реймента, вначале морской залив захватил юго-западную часть Африки (120 млн. лет назад), через 10 млн. лет он достиг ее центральной части. Море заполнило узкую зону грабена, прорезавшего Африку. 100 млн. лет назад уже существовала тогда еще очень узкая лента Атлантического океана.
Существенные изменения очертаний материков произошли в позднем мелу (100 - 80 млн. лет назад). Южная Америка поворачивается относительно Африки и перемещается на запад к своему современному местоположению. Индия отдаляется от Африки к северу, по направлению к Азии. Австралия и Антарктида испытывают перемещения к югу и юго-востоку от Африки. Африканский континент незначительно повернулся и несколько переместился на север до соприкосновения с Европой, отделяясь от нее Средиземным морем.
Первоначальные перемещения, вероятно, столь уж значительны. Атлантика, например, расширилась лишь на четверть от нынешней ширины. Тем не менее в мезозое была сформирована значительная часть дна Атлантического и Индийского океанов.
Геологические события в мезозойских геосинклиналях развивались в тесной связи с процессами, охватившими платформы. Как правило, геосинклинали располагались по периферии платформ в виде линейных прогибов. Мезозойские геосинклинальные области охватывали запад Америки, восток Сибири, Индокитай, Средиземноморье. Здесь формировались мощные толщи осадочных и магматических пород. В конце мезозоя активные тектонические движения мезозойской эпохи складчатости (ларамийская фаза) смяли пластичные осадочные породы геосинклиналей в складки, выдавили их вверх, образовав горные хребты. В результате многие геосинклинали заканчивают свое развитие, в их пределах устанавливается тектонический режим платформенного типа (рис. 16). Однако назвать их платформами можно лишь условно; правильнее говорить "пара-платформы" или "квазиплатформы", т. е. близкие к платформам. Типичный платформенный режим здесь еще не наступил. До сих пор происходит сглаживание, нивелировка горного рельефа, которое может продолжаться несколько миллионов лет. В ряде геосинклиналей (Андийская, Альпийско-Гималайская, Восточно-Азиатская) тектонические движения конца мезозоя не привели к смене режима, эти области земного шара продолжают свое геосинклинальное развитие и в кайнозое, хотя сокращаются в размерах.

Рис. 16. Схема современного положения мезозоид. 1 - параплатформы: 1 - Кордильерская, 2 - Верхояно-Колымская, 3 - Дальневосточная, 4 - Индокитайская; 2 - области более ранней консолидации; 3 - передовые прогибы: А - Предкордильерский, Б - Предверхоянский; 4 - срединные массивы - устойчивые области внутри мезозоид: а - Колымский, б - Омолонский, в - Охотский, г - Восточно-Чукотский, д - Индосинийский; 5 - Монголо-Охотский пояс
Интенсивность тектонических движений мезозойского времени была настолько велика, что горообразовательные процессы захватывают даже некоторые платформенные территории. Возникают так называемые области эпиплатформенной активизации, или эпиплатформенного орогенеза. Примером этого может служить Монголо-Охотский пояс, вовлеченный в активные горообразовательные движения в конце мезозоя.
В зоне сочленения мезозоид и древних платформ закладываются передовые прогибы. Платформы испытывают еще более четкую дифференциацию на области поднятий и области прогибаний.
Теплый мезозой. Мезозойская эра характеризовалась сравнительно теплым климатом. Исчезают ледники, которые существовали на Гондване в позднем палеозое.
Триасовый период был временем широкого развития засушливого климата. Пустыни и полупустыни охватывали Европу и Северную Америку. Районы Сибири и Индокитая обладали более влажным тропическим и субтропическим климатом.
По мере развития морских трансгрессий происходит глобальное смягчение климатических условий. В юре на значительных территориях, по-видимому, господствовала теплая, влажная обстановка. Тепло было даже в современной Арктике. Температура воды в районах современных Великобритании и Дании в ранней юре достигала +21°C, а в позднеюрскую эпоху она увеличилась до +28°C. Такие точные данные о палеотемпературах морских вод получены Г. Юри по соотношению изотопов кислорода в раковинках ископаемых моллюсков. Засушливые климатические зоны в юрском периоде сохранялись в Северной Америке, Южной Америке и Африке.
В течение мелового периода неоднократно происходили колебания климата, однако в общем сохранялись сравнительно теплые условия. Температура поверхностных вод в районах современной Арктики в то время равнялась +14°C, а на экваторе (океан Тетис) была даже выше современной. Общее смягчение климата, наметившееся еще в юрское время, привело к тому, что в позднем мелу практически не существовало пустынь в современном их понимании. Аридные (пустынные) зоны напоминали бы нам современную тропическую саванну с галерейными лесами и пышными зелеными оазисами в долинах рек.
Благоприятные климатические условия мезозоя способствовали бурному развитию органической жизни как на земле, так и в океанах. На континентах широко распространяются голосеменные растения, оттесняя плауновые, папоротниковые и хвощи. Развиваются хвойные, гингковые и беннетитовые растения, из которых до наших дней наиболее полно сохранились представители хвойных. В меловое время заметную роль начинают играть уже покрытосеменные растения, характеризующиеся двойным оплодотворением, возникновением завязи и плодов, развитием сложной проводящей ткани и вегетативными органами.
В мезозойских морях и океанах широкое развитие получила фауна беспозвоночных, которая взята за основу при расчленении мезозойских отложений. Преобладали аммониты со сложнопостроенной перегородочной линией и выразительной в скульптурном отношении спирально закрученной раковинкой (рис. 17, а, б), двустворки (пластинчато-жаберные моллюски), шестилучевые и восьмилучевые кораллы, губки, простейшие. Широко распространяются морские ежи - высокоорганизованные представители иглокожих, обладающие кровеносной и нервной системами. Тело животного было заключено в твердый известковый панцирь, защищенный тонкими известковыми иглами (рис. 17, в).

Рис. 17. Представители характерной мезозойской фауны. Головоногие моллюски, отряд аммоноидей: Туррилитгс (а), Паркинсония (б): известковый панцирь морского ежа, на пластинках видны точки прикрепления игл (в)
Наряду с обильной фауной беспозвоночных в мезозое довольно разнообразно представлены и позвоночные. В морях и океанах сокращается количество хрящевых рыб, их сменяют костистые рыбы. На суше в триасе появляются первые млекопитающие - мелкие животные размером в крысу. По особенностям строения зубного аппарата этих существ ученые заключили, что это были землеройки. Юрский период знаменуется развитием пернатых птиц.
Безраздельное господство среди животных захватывают рептилии, замечательно приспособившиеся к жизни на земле, в небе и морях. Такое обилие рептилий не встречалось ранее и вряд ли встретится в будущем. Остатки рептилий в ископаемом виде обнаружены на всех материках Гондваны. Показательны в этом отношении листрозаурусы - пресмыкающиеся, жившие в поздней перми и триасе (200 - 240 млн. лет назад). Это были приземистые животные, величиной с собаку (от 60 до 120 см). Массивное тело покоилось на коротких крепких ногах. Передняя челюсть на конце характерно изогнута книзу, все зубы вдавлены внутрь, и только два небольших клыка торчали наружу по обе стороны головы. Обитали они вблизи рек и озер субтропических и тропических областей. Находки листрозаурусов в Америке, Африке, Индии, Австралии и даже Антарктиде лишний раз подтверждают прежнее единство этих материков.
Среди мезозойских рептилий поражают своими размерами и необычными формами динозавры - самые крупные из сухопутных животных, населявших когда-либо нашу планету. Познакомимся поближе с этими любопытными животными, с их расцветом и вымиранием.
Пустыня Гоби и динозавры. В конце палеозоя среди рептилий были широко распространены сравнительно мелкие ящеры - текодонты, явившиеся родоначальниками динозавров. В триасе представители текодонтов приспосабливаются к передвижению на двух задних ногах. Поскольку у рептилий зрение - главный орган чувств, то высоко поднятая голова была особенно удобна для ориентировки среди густой травянистой растительности. Это дало определенные преимущества двуногим текодонтам, которые начинают доминировать среди животных.
Очаг возникновения динозавров, по мнению некоторых ученых, располагался в пустыне Гоби и в соседних районах Центральной Азии. Эти территории в течение последних 200 - 250 млн. лет не подвергались нашествию моря, что создавало благоприятные условия для стабильного развития наземной фауны. Здесь и по сей день часто находят остатки некогда живших гигантов. В Китае, например, с глубокой древности существовали добытчики ископаемых костей и зубов животных как лекарственного сырья. Из костей делались магические порошки, якобы лечившие от всех болезней. На Амуре, по данным академика Ю. А. Орлова, трубчатые позвонки динозавров рыбаки применяли для изготовления грузил. Понимая важность подобных находок для науки, Петр I в свое время издал указ, который гласил: "...Ежели кто найдет в земле или в воде старые вещи, а именно: каменья необыкновенные, кости человеческие или скотские, рыбьи или птичьи, не такия, какия у нас ныне есть, или и такие, да зело велики или малы перед обыкновенными... також бы приносили, за что давана будет довольная дача" (из указа Петра I, 13.XI.1718).
Наступление динозавров началось из Центральной Азии. Двигаясь на юг, динозавры заселяли территории Индокитая и Австралии; через Восточную Сибирь они проникли на Аляску, а оттуда в Америку; на западе перед ними открывались просторы Европы, Средней Азии и Аравии, откуда они проникли в Африку и освоили ее.
Динозавры имели сравнительно легкий скелет, что облегчало существование этих гигантов. Наиболее крупных размеров достигали травоядные динозавры. Известны диплодоки и бронтозавры длиной до 25 - 30 м и массой до 30 - 35 т. Но все рекорды побил сверхгигантский динозавр, кости которого были обнаружены американцем Д. Дженсеном в 1979 г. в горных районах штата Колорадо. Палеонтологом была выкопана лопаточная кость животного длиной в 3 м. По этой находке восстановлен облик динозавра: длина его тела 27 м, а живая масса 80 тонн! Гиганта окрестили "ультразавром". По-видимому, эти животные вели полуназемный образ жизни, проводя значительное время в воде подобно современным бегемотам. На это указывают облегченные шейные позвонки, полые кости скелета, тонкие слабые зубы, пригодные для поедания мягкой водной растительности, далеко сдвинутые назад глаза и носовые отверстия. Характерно в этом отношении строение черепа утконосого динозавра зауролофа (рис. 18, а). На затылке животного имеется "шпиль", образованный плотно сомкнутыми носовыми костями. Предполагают, что от переднего конца морды до конца "шпиля" по покрову кожи, одевавшей шпиль, проходил воздухоносный ход. Благодаря такому приспособлению, зауролоф мог полностью погружаться под воду в поисках пищи или спасаясь от врагов. Зауролоф был также приспособлен к плаванию, для чего мог использовать свой высокий и уплощенный хвост, кроме того, передние укороченные лапы были снабжены перепонками. Челюсти животного несли параллельные ряды листовидных зубов, число которых достигало 1000. Зубы были хорошо приспособлены к срезанию растительности, но быстро изнашивались, поэтому рост их происходил на протяжении всей жизни.

Рис. 18. Травоядные динозавры: а - населявший района Центральной Азии в позднемеловое время, реконструкция К. К. Флерова, б - игуанодон, реконструкция В. Свинтона (1958)
Полуназемные динозавры передвигались на четырех (диплодок) и на двух (зауролоф) ногах. Наземные же динозавры чаще всего были двуногими животными. Однако они также предпочитали влажные, богатые растительностью места. Одним из представителей наземных травоядных динозавров является игуанодон (рис. 18, б), достигавший 10 м в высоту. Животное имело трехпалые задние ноги с перепайками, что говорит о приспособленности игуанодонов к хождению по топким болотам и илистому дну озер и рек. "Батарея" зубов животного напоминала коренные зубы нынешних лошадей; конец морды был одет большим роговым чехлом, как у черепах и птиц. Своеобразный, клюв использовался при объедании листвы и побегов кустов и деревьев, а также для защиты. Игуанодон, имея сильную челюстную мускулатуру, мог наносить клювом опасные укусы и рваные раны, отбиваясь от хищников.
Страшными и мощными хищниками мезозойского времени были тарбозавры и тиранозавры (рис. 19), населявшие планету в меловом периоде. Встреча с такими чудовищами не предвещала ничего хорошего. Передвигались они на двух задних ногах, опираясь на сильный хвост. Укороченные передние конечности служили, вероятно, только для держания и разрывания пищи. Несмотря на огромный рост (до 12 м), тарбозавры и тиранозавры могли довольно быстро передвигаться. Возможно, что, используя хвост как балансир, эти динозавры даже проворно бегали, охотясь за травоядными животными. Мощные челюсти, усаженные острыми кинжальными зубами, делали этих животных непобедимыми. Динозавры размножались, откладывая яйца. Они зарывали их в горячий песок, как это делают современные черепахи и крокодилы (рис. 20). Некоторые представители динозавров вели стадный образ жизни. Американский палеонтолог Дж. Остром, изучая следы динозавров в Техасе и Коннектикуте, обратил внимание на то, что следы часто тянутся параллельной цепочкой. Похожие цепочки следов были обнаружены совсем недавно туркменскими геологами в Средней Азии на так называемой Ходжапильской площадке хр. Кугитанг. Осадочные породы позднеюрского возраста сохранили до наших дней почти полторы тысячи отпечатков тридцати пяти доисторических чудовищ, самый крупный след имеет в длину 80 см, самый маленький - 27 см. Обилие следов породило вначале предположение о переселении животных. Однако более внимательное изучение отпечатков отклонило эту гипотезу, ибо их трассы часто пересекались, расходились в разные стороны. Предполагается, что некоторые двуногие динозавры были довольно общительными животными и предпочитали жить и передвигаться небольшими группами. Так было легче отбиваться от хищников или же охотиться.

Рис. 19. Хищный тиранозавр, населявший Азию в меловом периоде. Реконструкция В. Свинтона (1964)
Постепенно рептилии приспосабливаются и к полету, захватывая главенство в воздухе. Предками летающих ящеров (птерозавров) также были текодонты, некоторые их представители обладали способностью к планированию и парашютированию, ведя древесный образ жизни. Не так давно в отрогах Туркестанского хребта был обнаружен возможный предок птерозавров, получивший название длинночешуйника необыкновенного. Животное величиной с мышь было покрыто чешуйками, напоминающими зачаточные перья. Вдоль спины располагались очень длинные чешуи, на концах они расширялись, образуя тонкостенные мешочки, которые и выполняли роль парашютиков, замедляя падение. Длинночешуйник обитал среди деревьев и питался насекомыми. Другие древесные рептилии присособились к планированию с помощью эластичной кожистой пленки, натянутой между конечностями и туловищем. От этих древесных рептилий, очевидно, и произошли крупные летающие ящеры (рамфоринхи, птеродактили и птеранодоны). Рамфоринхи имели длинный хвост, пятипалые задние ноги и длинные узкие крылья с размахом до 1 м. Челюсти были усажены острыми зубами. Тело животного покрывала густая шерсть длиной 4 - 7 мм, на перепончатых крыльях торчали лишь отдельные короткие волоски. Питались рамфоринхи рыбой и насекомыми. Птеродактили и птеранодоны в отличие от рамфоринхов обладали лишь зачаточным хвостом и широкими крыльями, размах которых достигал 8 м.

Рис. 20. Протоцератопсы - сравнительно небольшие растительноядные динозавры позднего мела. Реконструкция Ю. А. Орлова (1968)
Около десяти лет назад появилось сообщение о необычной находке в Национальном парке "Биг-Бенд" (штат Техас). Студент-палеонтолог Д. Лаусон обнаружил при раскопках кости птеродактилей, имевших размах крыльев более 17 м.
Древообитающие текодонты были, вероятно, и прародителями птиц. Появление последних приходится на юрский период. Птицы имеют очень много общего с рептилиями в строении черепа и скелета, поэтому их иногда называют "возвеличенными пресмыкающимися". Однако у птиц более совершенны кровеносная, нервная системы и органы чувств по сравнению с рептилиями. Большой объем мозга, гораздо лучше развитая "психика", их активность, забота о потомстве, координированный и совершенный полет - все это позволило им расселиться по всему земному шару, занять различные климатические области от полюсов до экватора и выжить в борьбе за существование, чего нельзя сказать о рамфоринхах и птеродактилях.
Начиная с триасового периода, некоторые рептилии приспосабливаются к морскому образу жизни. Наиболее совершенными представителями водных рептилий были, по-видимому, ихтиозавры. По внешнему виду они напоминали дельфинов, но с более вытянутой мордой. Длина животного составляла 5 - 6 м. Передние и задние ноги ихтиозавров преобразовались в ласты, причем передние были развиты значительно лучше задних. Ихтиозавры были живородящими животными, что отличает их от других рептилий. Об этом свидетельствуют находки захороненных животных с остатками детенышей внутри. По мнению Ю. А. Орлова, питание ихтиозавров в утробе матери происходило за счет материнского организма, как у млекопитающих. Все это говорит о более совершенном организме ихтиозавров по сравнению с остальными пресмыкающимися.
Другими обитателями мезозойских морей и современниками ихтиозавров были мозазавры, плезиозавры, плиозавры, нотозавры и плакодонты. Представители первых двух родов были наиболее приспособленными животными к жизни в морской среде. Мощные ласты позволяли им довольно быстро плавать, уступая, однако, в скорости ихтиозаврам. Небольшая голова, вооруженная пастью с острыми коническими зубами, на длинной "лебединой" шее помогала животным хватать проворных морских рыбешек. Наиболее крупные особи достигали в длину 13 м (эласмозавр).
Мезозой с полным правом можно назвать веком гигантских рептилий. Однако в конце мелового периода они практически полностью вымирают. Это произошло не молниеносно, но за довольно короткий в геологическом отношении период времени - около одного миллиона лет. Ничего подобного не случилось ни с рыбами, ни с примитивными млекопитающими, ни с амфибиями, ни с птицами, ни с растительным миром. Причины такого "мгновенного" исчезновения динозавров до сих пор еще не установлены, хотя имеется множество предположений.
Гибель гигантов. Наиболее распространенным объяснением причин вымирания динозавров было изменение климатических условий на Земле. Тектонические движения мезозойского тектогенеза, перемещение континентов приводили к изменению рельефа материков, сокращению площадей болот и низменностей. Климат становился более суровым и засушливым. Растительность приобрела сухой и жесткий характер, зубы же растительноядных динозавров были приспособлены к мягкой и сочной пище. Сокращение видов травоядных рептилий привело к гибели и хищников. Кроме того, динозавры, не обладая надежным волосяным покровом, не смогли наладить терморегуляцию организма, что также способствовало их вымиранию. Однако, несмотря на возможное изменение климата в сторону похолодания, на Земле все же существовали и теплые тропические зоны, где динозавры могли бы найти благоприятные условия для своего развития.
Большой интерес вызвала гипотеза о том, что гибель динозавров явилась следствием увеличения мутаций в организме. Причиной этого могла быть сильная радиация, нарушившая производство потомства. Американские ученые подсчитали, что каждые 10 млн. лет наша планета может получать дозу космических лучей, превышающую нормальную в семь тысяч раз. Такие условия создаются, например, при взрыве сверхновых звезд. Если это так, то почему бомбардировка жестким излучением оказалась смертельной только для динозавров? Кроме того, специальные исследования ископаемых костей этих животных не показали их повышенную радиоактивность.
Некоторые ученые видели причины гибели гигантов животного мира в несоответствии между объемом мозга и массой тела, а также в вирусной эпидемии, поразившей динозавров. Профессор Г. Эрбен из Бернского университета предположил, что на последней стадии существования динозавры откладывали яйца с очень толстой скорлупой. Детеныши могли пробить ее с трудом, и далеко не всегда, в результате рождаемость динозавров резко снизилась.
Московский геолог В. Б. Нейман связывает гибель гигантов с возрастанием силы тяжести на Земле, они оказались "раздавленными собственной массой". Исследователь приводит некоторые данные, показывающие увеличение силы тяжести и в наши дни. Предположение само по себе не лишено смысла. Однако среди динозавров были и не такие уж исполины. Морские рептилии, например, по размерам и массе уступали современным китам. По-видимому, что-то роковое было в самой "конструкции" организма у динозавров.
Существует еще одна, как нам кажется, наиболее правдоподобная гипотеза, объясняющая гибель гигантов, основанная на изменении микроэлементов в окружающей среде. В конце мезозоя значительную роль в строении земной коры начинают играть континенты. Эрозионные процессы, охватившие их, выносят в атмосферу и гидросферу новые соли и микроэлементы, до этого активно не участвовавшие в круговороте веществ. Появление новых микроэлементов, изменение солевого состава водоемов могло коренным образом повлиять на судьбу динозавров.
Исследования последних лет, например, установили, что нарушение баланса микроэлементов пагубно сказывается на организме человека и животных: падение количества натрия в крови вызывает истощение нервной системы, быструю утомляемость; дефицит железа способствует развитию малокровия; уменьшение кальция - важнейшего элемента скелета - приводит к сокращению размеров животных; недостача йода влечет за собой увеличение щитовидной железы; нехватка фтора - болезнь зубов.
Сильное влияние на организм оказывает стронций. Повышенное содержание его в почвах приводит к тому, что стронций заменяет кальций в костной ткани. Сравнительно быстро он удаляется из организма и кости становятся пористыми - развиваются болезни скелета, резко возрастает ломкость костей.
Не менее вредное влияние на организмы оказывает селен. Из почвы он попадает в растения, которые охотно поедаются животными. У них развивается "щелочная болезнь": нарушается работа сердца, поражаются суставы, выпадает волосяной покров, размягчаются рога и копыта. Мясо таких животных ядовито. Другие примеры вредного влияния микроэлементов на организмы: недостаток кобальта вызывает эндемические заболевания, авитаминоз, недостаток меди - анемию.
Возможно, что в конце мезозоя произошло выщелачивание из почв и горных пород каких-то особо вредных для динозавров микроэлементов. Включившись в круговорот веществ, они попадали в организмы животных, что способствовало развитию тяжелых недугов, явившихся причиной гибели самых крупных существ, некогда обитавших на Земле. Это предположение подтверждается интересными данными, недавно полученными физиком Калифорнийского университета Л. Альваресом и его сыном - геологом У. Альваресом. Ими было доказано, что в пограничном слое между меловыми и палеогеновыми отложениями на уровне примерно 65 млн. лет резко увеличивается содержание тяжелого металла иридия в 30, 160 и даже в 200 раз. Содержания же иридия, как и других тяжелых металлов, ядовиты для животных. "Иридиевую аномалию" объясняют падением на Землю кометы, астероида или крупного метеорита, не только доставившего избыток этого ядовитого элемента, но и резко нарушившего сложившиеся условия жизни на ее поверхности и в атмосфере нашей планеты.
Британский палеонтолог профессор Т. Свейн объясняет гибель травоядных динозавров отравлением от растительности. Эти ящеры съедали за день огромное количество зеленой массы. Чтобы поддержать жизнь в своем гигантском теле, они должны были безостановочно глотать мягкие водоросли и листву - на жевание у них просто не оставалось времени. В конце мезозойской эры - этого рокового для динозавров рубежа - появляются покрытосеменные растения, содержащие танин, алкалоиды (стрихнин и морфин). Попадая в большом количестве в организм, они отравляли травоядных динозавров, что в свою очередь и привело к исчезновению хищных.
Есть и другие гипотезы. Одна из них связывает гибель гигантов с усилением напряженности геоэлектрических и атмосферно-электрических полей. Активизация в конце мезозоя тектонической жизни (землетрясения; вулканы) приводит и к активизации электрических процессов в атмосфере. Участившиеся электрические удары и молнии поражают беззащитных великанов.
Кайнозойская история
Последний этап геологического развития Земли, приведший ее к современному состоянию, приходится на кайнозойскую эру. Продолжительность его самая незначительная по сравнению с предыдущими этапами - около 70 млн. лет. Однако этого достаточно, чтобы материки резко изменили свои очертания и заняли привычные места на географических картах; значительно выросли Атлантический и Индийский океаны; существенно изменились растительность и органический мир; наконец, появился человек и достиг, большого могущества.
Кайнозойская эра делится на три периода: палеогеновый, неогеновый и антропогеновый (или четвертичный). Каждый из них в своем составе имеет эпохи и века.
"Расползание" материаков продолжается. В начале кайнозойской эры, несмотря на то что уже произошел раскол Лавразии и Гондваны и отдельные материковые глыбы литосферы начали "расползаться", все-таки мы застали бы еще довольно крупные участки суши, охватывавшие иногда несколько современных континентов. Единый материк, возможно, составляли Индия, Австралия и Антарктида; Южная Америка еще имела точки соприкосновения с Африкой. Между Европой и Северной Америкой существовала лишь узкая полоска воды - зачаток будущей Северной Атлантики. Тектонические движения кайнозойского времени существенно изменили этот лик Земли.
В начале палеогенового периода развивается крупная морская трансгрессия, захватившая эпигерцинские платформы северного полушария, затопившая юг Восточно-Европейской платформы, Аравию и север Африки. Северо-Американская и Сибирская платформы оставались сушей.
В морях накапливались мелководные осадки (пески, глинистые и карбонатные илы), а на суше - речные и озерные осадки. Наступление моря шло в основном со стороны океана Тетиса, где продолжала формироваться крупная Альпийско-Гималайская геосинклинальная область. Для нее было характерно интенсивное прогибание земной коры и накопление глинистых, песчаных и известковых толщ, образовавших в ряде мест специфическую флишевую формацию1 палеогена мощностью до 5 км. Геосинклинальное развитие испытывает и Восточно-Азиатская зона, включающая в себя крайнее побережье Тихого океана. Аналогичные условия накопления осадков сохраняются практически в течение всего неогена и антропогена.
1 (Формация - закономерное сочленение пород, объединенных общностью тектонических условий образования. Флиш - частое ритмичное чередование пластов известняка, мергеля, глин, песчаника)
Морское осадконакопление присуще геосинклиналям, отчасти эпигерцинским платформам (Западная Сибирь, Средняя Азия, Предкавказье). Древние же платформы не перекрывались, лишь их периферические области иногда испытывали трансгрессию моря. Обширные территории этих платформ служили областями сноса обломочного материала в прилегающие водные бассейны геосинклиналей, поэтому осадконакопление на них шло преимущественно в изолированных озерных или болотных водоемах или же в пойме речных долин. Однако и в этом случае создавались иногда благоприятные условия для накопления мощных толщ осадков. Наиболее значительные мощности кайнозойских отложений установлены в долине р. Миссисипи, активно развивавшейся на Северо-Американской платформе в течение всей кайнозойской эры. Общая мощность русловых и дельтовых осадков, накопившихся в неоген-антропогеновое время, составляет около 20 км. Образование такой толщи связано с интенсивным прогибанием узкого грабенообразного желоба, расположенного между двумя поднятиями платформы: Цинцинати и Озарк. Американские геологи рассматривают эту зону в качестве своеобразного геосинклинального трога.
В неогеновом периоде продолжается "расползание" материков. Особенно интенсивно формировалась Атлантика. Постепенный рост этого океана за последние 50 млн. лет фиксируется изменением возраста островов. Радиогеохронологическими методами было установлено, что близкие к суше острова имеют более древний возраст, чем острова, расположенные в центральной части океана. Так, острова Зеленого Мыса, Принсипи, Сан-Томе, Аннобон (вдоль западного побережья Африки) и Багамские, Фернанду-ди-Норонья (вдоль восточного побережья Северной и Южной Америки) имеют возраст 120 - 150 млн. лет. Острова Азорские, св. Елены, Гоф, Найтингейл, Бермуды намного моложе - они не старше 30 - 20 млн. лет. Наконец, острова Тристан-да-Кунья, Буве, Ян-Майен, лежащие почти на оси Атлантического океана, молоды - им не более 10 млн. лет. Учитывая возраст островов и их расстояние до материков, можно вычислить и скорость удаления Африки и Европы от Северной и Южной Америки. Она составит 2 - 6 см/год.
В палеогене уже произошло разделение Индии, Австралии и Антарктиды. Индийская глыба переместилась почти на 8 тыс. км к северу и в начале неогена вошла в соприкосновение с Азией. Австралия двигалась на северо-восток, поворачиваясь вокруг своей оси против часовой стрелки. Менее всего подвижны оказались Антарктида и Африка.
Великие Африканские разломы. Одновременно с перемещением континентальных блоков литосферы, очерченных глубинными разломами еще в начале мезозоя, в кайнозойской эре происходило заложение новых глубинных разломов, в ряде случаев приведших к изменению географии Земли.
Крупные глубинные разломы с активными вертикальными движениями по ним возникали на западе Северо-Американской платформы, в зоне сочленения с Кордильерами. Блоковые движения привели здесь к образованию горного рельефа (Скалистые горы) и излиянию базальтовых лав, покрывших многосотметровым плащом площадью свыше 500 000 км2.
Еще более грандиозные расколы перекроили территорию Аравии и Африки. В миоценовую эпоху система взаимно пересекающихся диагональных глубинных разломов образовала грабены-рифты Красного моря, Суэцкого и Аденского заливов. Узкие грабены, ориентированные в северо-западном и северо-восточном направлениях, отделили Аравийский полуостров от Африки. На протяжении последних 5 млн. лет края этих грабеноврифтов неумолимо отодвигаются в разные стороны, а образовавшееся пространство заполняется морской водой.
Земная кора Красного моря, Суэцкого и Аденского заливов имеет океаническое строение. Ученые считают, что образование подобных грабенов является начальным этапом формирования океанических впадин на теле Земли. Исследования, проведенные в придонных водах Красного моря, показали, что активные термические процессы, давшие, вероятно, толчок к образованию грабенов, протекают и в настоящее время. Придонные воды Красного моря нагреты до +61°C, их минерализация резко возрастает до 27%1 за счет повышенного содержания различных солей, а также цинка, золота, меди, железа, серебра, урана. Это указывает на прямую связь вод с глубинными недрами и на вынос оттуда различных элементов. Насыщенная минеральными солями придонная вода Красного моря не поднимается к поверхности, хотя сильно нагрета, а образует своеобразный "рассол", концентрирующийся на морском дне на глубине 2 - 2,5 км.
1 (Средняя минерализация вод Красного моря 4%, а температура 27°C)
По некоторым разломам Африки происходит активное излияние лав на дневную поверхность. В палеогене вспышка вулканической деятельности произошла в центральных и юго-западных районах Африки. Мощность базальтовых покровов палеогена достигает здесь 1,5 км.
В миоцене Восточная Африка "вспарывается" грандиозной системой глубинных разломов-рифтов. Начинаясь у нижнего течения р. Замбези, разломы тянутся к северу в субмеридиональном направлении. У оз. Ньяса они расчленяются, давая три ветви. Западная ветвь простирается в северо-западном направлении через озера Танганьика и Эдуарда и вскоре затухает, а восточная поворачивает на северо-восток и около южной оконечности п-ова Сомали выходит к Индийскому океану. Центральная ветвь трассируется в северном направлении через озера Рудольфа и Дофине, где снова делится на две ветви: короткая восточная подходит к побережью Аденского залива, а длинная западная - через Эфиопию проходит к Красному морю и далее к Мертвому морю, заканчиваясь у южного подножья горного хребта Тавр в Малой Азии. Строго говоря, в систему Великих Африканских разломов входят и грабены Красного моря, заливов Аденского и Суэцкого.
В рельефе местности разломы выражены узкими, длинными и глубокими грабенами, заполненными водой: это озера Рудольфа, Ньяса, Танганьика, Эдуард, Альберт и др. Часть разломов образует ступенчатые сбросы, погружающиеся в сторону Индийского океана. Образование разломов и их развитие сопровождались интенсивным излиянием преимущественно базальтовой лавы, которое продолжается и в антропогеновое время. Действующие вулканы Африки, как правило, "сидят" на Великих Африканских разломах, за исключением вулкана Камерун, связанного с Камерунским глубинным разломом.
Образование молодых грабенов-рифтов в зоне активных глубинных разломов отмечается ив некоторых других районах земного шара. Так, в Сибири образовался грабен оз. Байкал, амплитуда проседания которого составила около 1700 м. На западе Северной Америки по глубинному разлому произошло отделение Калифорнии от материка с образованием Калифорнийского залива. Горизонтальные движения по глубинному разлому Сан-Андреас, разделяющего Северную Америку и Калифорнию, происходят и по сей день, сопровождаясь сильными землетрясениями, со средней скоростью 5 - 6 см в год.
От Пиренеев до Гиндукуша. В неогеновое время жесткие плиты литосферы продолжают перемещаться и в ряде мест приходят в соприкосновение друг с другом. При этом мягкие осадочные породы сминаются с образованием горных хребтов. Индийская глыба, приблизившись к Азии, вызвала образование мощной: горной системы мира - Гималаев. Продвижение Африки к северу привело к "вздыбливанию" Средиземноморской системы гор - Атласа, Альп, Пиренеев и др. Ученые подсчитали, что Альпы до этого имели ширину 600 - 1200 км, а после сжатия сократились до 150 км. Процессы смятия мощных толщ осадочных пород геосинклиналей, разделявших в кайнозое жесткие глыбы платформ, проявлялись отдельными импульсами, которые выделяются как фазы альпийского диастрофизма. Суммарным результатом их явилось возникновение альпийских областей складчатости (рис. 21).

Рис. 21. Схема современного положения кайнозойских структур. 1 - области альпийской складчатости: 1 - Северо-Американская, или Береговые хребты, 2 - Южно-Американская, или Андийская, 3 - Средиземноморская, 4 - Понто-Иранская, 5 - Гималайская, 6 - Восточно-Азиатская, 7 - Индонезийская; 2 - области более ранней консолидации; 3 - основные области эпиплатформенной активизации: I - Скалистые горы, II - Тянь-Шань, III - Тибет, IV - Восточно-Африканская; 4 - передовые прогибы: А - Предальпийский, Б - Предкарпатский, В - Предкавказский, Г - Предкопетдагский, Д - Предрамирский, Е - Индский, Ж - Гангский, З - Предандийский, И - Месопотамский
Одна из грандиознейших областей альпийской складчатости протянулась от Пиренеев до Гиндукуша и далее до Гималаев на расстояние почти в 12000 км; при ширине до 1000 км. Она получила название Альпийско-Гималайского горно-складчатого пояса. Геосинклинальная история его еще не закончилась, эта область переживает в настоящее время орогенный, т. е. горообразовательный этап развития. Частые землетрясения, извержения вулканов свидетельствуют о том, что высокогорные страны мира, расположенные в Альпийско-Гималайском поясе (Альпы, Памир, Гималаи, Гиндукуш и др.), продолжают "расти", все выше поднимая над земной поверхностью свои снежные вершины. Такого же типа альпийская область известна в Южной Америке - это Анды.
На востоке Азии от Корякского нагорья до Новой Гвинеи, захватывая п-ов Камчатку, Курильские, Японские и многие другие острова, расположен Восточно-Азиатский пояс. Тектонические движения альпийской складчатости не привели здесь к ликвидации геосинклинального режима развития. Островные дуги выполняют роль внутригеосинклинальных поднятий. Эрозионные процессы сносят с их поверхности обломочный материал, который поступает в прилегающие окраинно-материковые моря (Японское, Восточно-Китайское, Южно-Китайское, Сулавеси и др.). Многочисленные вулканы извергают лавы. Все это накапливается в морях, являющихся внутригеосинклинальными прогибами. В их пределах мощность кайнозойских отложений осадочно-эффузивных образований превышает местами 10 - 15 км. Аналогичный режим развития испытывает: и сравнительно узкая зона Северной Америки, захватывающая Алеутские острова, береговые хребты Кордильер и п-ов Калифорнию.
Мощь тектонических движений кайнозойского времени была настолько велика, что вызвала горообразование даже в пределах платформенных областей. С такими областями эпиплатформенного орогенеза мы сталкивались уже в конце мезозоя, когда возник Монголо-Охотский пояс. На кайнозойском этапе развития масштабы образования областей эпиплатформенной активизации были более значительны. Омоложение горного рельефа произошло на Урале, Тянь-Шане, Алтае, в Забайкалье, Саянах и в других районах земного шара.
Между молодыми горными сооружениями альпийского возраста и существовавшими платформами закладываются передовые прогибы.
Легендарный океан Тетис. Бурные тектонические процессы, приведшие к перемещению континентов, к образованию мощнейших горных поясов, сильно изменили географию планеты: возникли новые океаны (Атлантический, Индийский) и исчезли прежние. Печальная участь постигла в этом отношении легендарный океан Тетит. В течение сотен миллионов лет этот океан, по-видимому, был одним из крупнейших океанических бассейнов планеты.
В неогене, когда молодые горообразовательные движения "набрали" силу, океан Тетис распался вначале на два неравных по величине морских бассейна: северный и южный. Узкая полоска земли, протягивавшаяся от Альп через Балканы, Крым, Кавказ в Центральный Иран и Афганистан, разграничивала эти моря. Более крупный южный бассейн сохранял связь с Мировым океаном. Северный оказался изолированным, растущие горные сооружения все больше отшнуровывали его от открытого океана. Возникло гигантское Сарматское море-озеро - Паратетис, простиравшееся на тысячи километров из внутренних районов Европы вдоль Предкавказья до Аравийского моря. В этом бассейне, перекрывавшем и юг Восточно-Европейской платформы, формировались песчано-глинистые осадки, органогенные известковые илы с богатой фауной моллюсков, фораминифер.
В плиоцене Паратетис расчленился на ряд изолированных бассейнов - сказывались продолжавшиеся горообразовательные процессы и рост горных сооружений. Растущие горы Кавказа и Копетдага вызывали перемещение водных масс Паратетиса на север. В дальнейшем, в позднем плиоцене, затапливается вся современная Прикаспийская низменность. "Языки" моря по долинам палеорек Волги, Камы, Урала проникают далеко над север. Плиоценовые моря были сильно опреснены, в них формировались исключительно песчано-глинистые отложения. Отступила и южная часть океана Тетис, имевшая связь с открытым океаном.
С течением времени горные сооружения Альп, Карпат, Кавказа, Копетдага принимают современные очертания, Они растут, увеличивают свои размеры, Воздымание захватывает и прилегающие территории, что сказывается и на оставшихся от океана Тетис морских бассейнах. Южный морской бассейн регрессирует все дальше на юг, постепенно уходя в Индийский океан, оставляя после себя лишь небольшие морские водоемы, соленые озера, заливы, которые в конце концов полностью пересыхают.
Паратетис испытывал все большую разобщенность: изолированность отдельных морских водоемов возрастает. Обособились Средиземное, Черное, Каспийское моря. В позднем плейстоцене образуется Аральское море-озеро. Периодическая связь между ними восстанавливается и обрывается вновь. Тогда еще не существовало Гибралтарского пролива, Босфора, и Дарданелл, Эгейского моря. Европа непосредственно соединялась в ряде мест с Африкой. Некоторые из возникших изолированных бассейнов периодически полностью пересыхали. Такая участь, по мнению швейцарского ученого К. Хсу, постигла Средиземное море. Отсутствие притока воды привело к тому, что это море постепенно усыхало, концентрация солей в нем возрастала все более и более.
По современным подсчетам ежегодно за счет испарения Средиземное море теряет около полутора тысяч кубических километров воды. И если закрыть Гибралтарский пролив, то примерно через тысячу лет море полностью высохнет. Поэтому ничего нет удивительного, если, оказавшись в районе Средиземного моря 6 - 7 млн. лет назад, мы вместо него обнаружили бы огромную равнину, лежавшую на 3 тыс. м ниже уровня Мирового океана. Солнечные лучи превратили эту гигантскую чашу в знойную пустыню, поверхность которой была покрыта не песком, а каменной солью, ангидритом и гипсом. Предполагают, что примерно 6 млн. лет назад Гибралтарский хребет, соединявший Европу и Африку, рухнул в результате сильного землетрясения и воды Атлантики невиданным каскадом хлынули, наполняя Средиземное море. Пропускная способность Гибралтарского водопада была в сто раз больше, чем у водопада Виктория, и в тысячу раз больше, чем у Ниагарского. В конце плиоцена заливы моря вторгались в долину Нила, поднявшись вверх по его течению на 1200 км.
Сложную историю испытывали и другие остаточные морские басейны океана Тетиса. В неогеновое и антропогеновое время Черное, Каспийское и Аральское моря неоднократно меняли свои очертания, периодически трансгрессируя на сушу. В антропогене возникла связь Черного моря со Средиземным через проливы Босфор и Дарданеллы, что спасло Черное море от усыхания. Наоборот, связь его с Каспием была окончательно нарушена. В настоящее время Каспийское и Аральское моря медленно, но неуклонно сокращают свою площадь. Вода, приносимая реками, не компенсирует испарения.
Таким образом, в неоген-антропогеновое время произошли окончательный распад и гибель океана Тетис. Теперь только пересыхающие внутриконтинентальные моря напоминают о некогда грандиозном океане, разделявшем Лавразию и Гондвану.
Оледенение в Европе. Равномерно теплый климат застали бы мы на земном шаре в палеогеновый период. Климатические зоны, являющиеся следствием контраста температур между полюсами и экватором, в то время были как бы стерты за счет более высоких температур в приполярных и полярных областях обоих полушарий. Зона тропического и субтропического климата захватывала не только экваториальный пояс, но и районы Европы, Кавказа, Предкавказья, Нижнего Поволжья. Здесь в отложениях палеогена и миоцена находят остатки теплолюбивой флоры: пальм, магнолий, лавра. Далее к северу проходил умеренный пояс, включая Гренландию и Шпицберген.
Умеренный климат был характерен и для районов Аляски и Анадырского края. Здесь росли дубовые, буковые и каштановые леса. По-видимому, полярной климатической области в северном полушарии тогда вообще не существовало. Температура воды Северного Ледовитого океана, по данным П. М. Борисова, достигала +14°C, а на экваторе она была даже намного выше, чем в наши дни. Мировой океан был теплым практически по всей толще воды. Температура донных осадков в олигоцене в экваториальной зоне составляла + 10°C, в миоцене 7°C, а в плиоцене она достигала уже современных значений 2°C.
В палеогене и начале неогена, вероятно, не существовало пустынь. Засушливые, аридные зоны, занимавшие территорию современной Туркмении, Монголии, Египта, Испании, больше напоминали африканскую саванну, чем пустыню.

Рис. 22. Изменение среднегодовой температуры в Западной Европе за последние 60 млн. лет. По П. Вольдштедту. Горизонтальный масштаб ангропогенового времени увеличен в 10 раз
Теплые климатические условия продолжались до конца миоценовой эпохи. Ничто, казалось бы, не предвещало суровых испытаний. Однако в конце миоцена началась одна из крупнейших в истории Земли депрессия мирового климата, кульминация которой наступила 45 - 60 тыс. лет назад.
За последние 60 млн. лет среднегодовая температура устойчиво падает (рис. 22). С эоцена по миоцен это происходило постепенно: по районам Западной Европы изменение среднегодовой температуры составило всего 7°C (с 22 по 15°C). За плиоценовую эпоху падение среднегодовой температуры составило уже 13°C (с 15 до 2°C). В антропогене наблюдается неоднократное "лихорадочное" чередование повышения и понижения среднегодовой температуры с общей тенденцией к ее снижению.
В антропогене мощное оледенение охватывает континенты северного полушария. В Европе центрами оледения явились Скандинавский полуостров и Альпы. Особенно большие следы оставил Скандинавский ледник. Ледниковый покров охватил территорию нынешних Великобритании, Центральной Европы, Урала и Валдая. Край ледника доходил до Волгограда, а по Днепру ледник спускался до 48° с. ш. Периоды наступления ледника сменялись эпохами потепления, во время которых ледник отступал. Кривая изменения среднегодовых температур в Европе в антропогеновое время имеет сильно изрезанный характер, что и соответствует чередованию ледниковых и межледниковых эпох. В Западной Европе насчитывают четыре эпохи максимального оледенения: гюнц (поздний плиоцен), миндель, рисе, вюрм; в европейской части Советского Союза - три: окскую, днепровскую и валдайскую. Максимум оледенения приходится на рисскую и днепровскую фазы.
Обширные оледенения захватили также азиатскую часть СССР и Северную Америку (рис. 23). Центры ледников располагались на Новой Земле, Таймыре, в Забайкалье и Гренландии. В Северной Америке ледник перекрыл всю северную половину континента, опускался по долине р. Миссисипи южнее 37° с. ш., заканчиваясь в 1500 км от линии тропика.
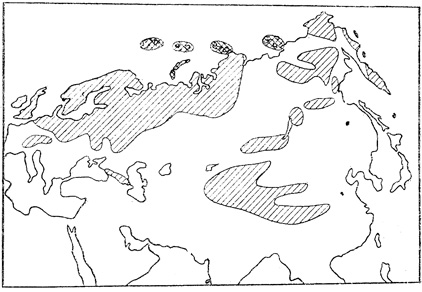

Рис. 23. Максимальное распространение льдов в северном полушарии в антропогене. По К. К. Маркову. 1 - современные области материкового льда; 2 - области четвертичного оледенения
Мощность четвертичных ледников предположительно достигала 3,5 км, а в районе Новой Земли 4 км. В это время в северном полушарии было сосредоточено до 67% площади континентальных ледников земного шара, тогда как ныне эта величина не превышает 16%. Последние примерно 10 тыс. лет приходятся на послеледниковую эпоху. Границы ледников в северном полушарии резко сократились (на 91%). Материковые льды остались в Гренландии и на некоторых островах Северного Ледовитого океана. Общая площадь их не превышает 2,3 млн. км2.
В ледниковые эпохи значительно усилилось планетарное похолодание, уменьшилось увлажнение материков, увеличилась континентальность. Оформились границы климатических зон, сохранивших свое положение и по сей день.
Существуют различные точки зрения, объясняющие возникновение четвертичных ледников в Северном полушарии. Это могло произойти в силу уменьшения притока тепла при изменении угла наклона земной оси к плоскости эклиптики. По подсчетам шведского ученого С. Аррениуса (1902 г.), увеличение содержания углекислоты в воздухе в три раза должно бы повысить температуру поверхности Земли на 8 - 9°C. При этом содержание углекислоты в атмосфере ставится в зависимость от вулканической активности, поэтому снижение ее приводит к похолоданию климата, и наоборот, усиление деятельности вулканов может вызвать потепление климата. По данным же английского климатолога Ч. Брукса, повышения температуры на поверхности Земли всего на 1°C оказалось бы достаточным, чтобы привести весь ледяной покров Антарктики в неустойчивое состояние.
В последнее время высказывается гипотеза, согласно которой периодическое похолодание или потепление происходит за счет изменения орбиты движения Земли вокруг Солнца. Примерно каждые 100 тыс. лет конфигурация орбиты меняется от эллиптической до круговой и снова до эллиптической. Ледниковый период наступает при круговой орбите, когда Земля равномерно и в течение длительного времени удалена от Солнца. Изменение орбиты в сторону эллиптической позволяет земному шару близко подходить к нашему светилу и прогреваться.
Торжество млекопитающих. Благоприятные климатические условия палеогена и миоцена способствовали бурному развитию растительности и животного мира. Господство среди растений захватывают покрытосеменные. В Европе и Азии обособляется гелинденская флора с тропическими растениями (пальмы, кипарисовидные, папоротниковые), в северной части Европы распространяется листопадная (гренландская) флора - бук, дуб, платан, хвойные. С течением времени в связи с общим похолоданием гелинденская флора переходит во флору умеренного пояса (тургайскую), представленную березой, кленом, ольхой. В антропогене, в ледниковый период, происходит сокращение лесов в северном полушарии, на смену которым приходит тундровая растительность: мхи, лишайники, карликовые деревья.
В теплых морях и океанах среди беспозвоночных животных, составляющих основу при расчленении кайнозойских толщ, распространены моллюски (пелециподы и гастроподы), кораллы (рис. 24), простейшие (нуммулиты, фораминиферы, радиолярии). Последние часто становятся породообразующими организмами.

Рис. 24. Характерные представители кайнозойской фауны. Пластинчато-жаберные моллюски: Спондилус (а), Пектен денудатус (б), Кардиум фиттони (в), современный восьмилучевой коралл Тубипора (г)
К неогену количество простейших резко сокращается. В умеренных широтах развиваются диатомовые водоросли, послужившие материалом для образования опок и трепелов - характерных осадочных пород палеогена. Из хордовых широко представлены костичные рыбы, птицы, земноводные (лягушки, саламандры), пресмыкающиеся (черепахи, крокодилы, чешуйчатые).
В кайнозое активно развиваются млекопитающие, оказавшиеся наиболее жизнеспособными. Они приспосабливаются к жизни на деревьях (белки, соболи, куницы, некоторые приматы), в морях (китообразные и ластоногие), к полету в воздухе (летучие мыши). Кайнозой - время торжества млекопитающих.
Для палеогена характерны клоачные, сумчатые и примитивные плацентарные млекопитающие, насекомоядные, лошадиные, хоботные, древние хищники. В неогене расцвета достигают плацентарные млекопитающие: хищные, травоядные и приматы. Появляются медведи, мастодонты, носороги, быки, человекообразные обезьяны, а в конце неогена - слоны, гиппопотамы, гиппарионы, лошади (гиппарионовая фауна).
Важные изменения происходят в антропогене. Вымирают многие представители гиппарионовой фауны, на смену им приходят животные, приспособившиеся к жизни в холодных тундрах и лесостепях, возникших в результате плейстоценовых оледенений в Европе. Это большерогие и благородные олени, волосатые носороги, зубры, мамонты, туры, медведи.
Выдающимся событием антропогенового периода является появление человека, существенно повлиявшего на дальнейшую эволюцию жизни на Земле. По мнению американских ученых П. Мартина и Г. Краптца, исчезновение крупных млекопитающих (мамонтов, бизонов, носорогов) и крупных птиц (моа, динорнис) в четвертичное время явилось делом рук человека, - нарушившего экологическое равновесие до такой степени, что конечный результат оказался весьма плачевным. Подтверждением этого явилось обнаружение в последние годы в районе с. Межирич на Украине огромного скопления костей мамонтов. На площадке в четверть футбольного поля находились останки 204 гигантских зверей, а также множество костей древних лошадей, оленей, бизонов. Кости мамонтов использовались людьми каменного века для сооружения "домов", которые служили местом культовых охотничьих обрядов. По-видимому, культура палеолитических охотников с односторонней ориентацией на крупных травоядных в значительной степени способствовала исчезновению гигантов.
Вымиранию гигантских птиц на Мадагаскаре способствовали земледельцы. Такое же роковое влияние, по мнению английского ученого Клоудсей-Момпсона, оказала римская цивилизация на развитие животного и растительного мира Сахары. Корчевание лесов, земледелие и животноводство привело 4 тыс. лет назад к изменению климата Северной Сахары в сторону его засушливости. Исчезли водоемы, леса и возникли пустыни. Еще 6 тыс. лет назад на месте крупнейшей современной пустыни мира обитали влаголюбивые гиппопотамы!
Человека называют царем природы, но свое "царствование" он должен осуществлять разумно и мудро, не забывая, что сам он - часть этой природы.
Две секунды человечества. История человека насчитывает около 2 млн. лет, а в нашем волшебном кинофильме - это всего две секунды! Однако их оказалось достаточно для захвата человеком безраздельного господства на Земле и освоения околоземного космического пространства. Приостановим скорость волшебного кино, и в замедленной съемке посмотрим появление и развитие человека.
Появлению человека предшествовала длительная эволюция приматов, напоминавших по внешнему виду обезьян и явившихся общими предками современных человекообразных обезьян и человека. Профессор антропологии Гарвардского университета У. Хауэльс в качестве такого общего прародителя называет дриопитека, обитавшего около 20 млн. лет назад в Европе, Индии, Китае и напоминавшего человекообразную обезьяну. Из группы дриопитека примерно 12 млн. лет назад выделяется рамапитек - первый предок человека. По внешнему виду он больше походил на обезьян, но уже имел некоторые признаки человека.
Первый гоминид - австралопитек был обнаружен в Южной Африке в 1924 г. Он жил 6 - 1,5 млн. лет назад. Австралопитек ("южная обезьяна") уже миновал основную веху в своей эволюции. Подобно человеку австралопитек мог быстро передвигаться по открытой местности, опираясь на стопу при прямом торсе. Рост его составлял 90 - 120 см. Вместе с остатками австралопитека в отложениях близ оз. Рудольфа в Кении обнаружены примитивные каменные орудия: речные гальки, заостренные 5 - 8 сколами. Возраст этих орудий 2,6 млн. лет. В более молодых наслоениях, возникших 1,5 млн. лет назад, близ оз. Баринчо (Кения) обнаружены следы древних костров, куски спеченной глины, кремневые кресала и кости самих австралопитеков. Все это говорит о сравнительно высоком развитии "южных обезьян", хотя объем мозга у них составлял всего 400 см3, т. е. в четыре раза меньше, чем у современных людей. Следующим важным шагом на пути развития человека является появление "человека выпрямленного" - Homo erectus около 1 млн. лет назад. С этого момента появляется новый род у приматов - "Homo", констатирующий новые отличительные качества и знаменующий появление человека. Ряд антропологов (например, Р. Лики) считают первым человеком уже некоторых австралопитеков - "человека умелого" - Homo habitis. "Человек выпрямленный" уже пользовался огнем и создал первые ручные рубила (аббевильская культура). 250 тыс. лет до н. э. Европу населял "человек разумный" ранний Homo sapiens. Это был наиболее древний представитель человека современного вида. Режущие орудия "человека разумного" были единообразны, хорошо обработаны, им придавалась форма, близкая к простейшим геометрическим формам (ашельская культура).
В период 150000 - 35000 лет Европу, Африку, Азию заселяли палеоантропы (неандертальцы) - более высокоорганизованные представители рода Homo. Неандертальцами создаются разнообразные кремневые орудия, изготовлявшиеся с применением ретуши, что знаменовало собой определенный прогресс (мустьерская культура).
Дальнейшая история эволюции человека не совсем ясна. По мнению одних ученых, неандертальцы совершенствовались и далее, другие же считают, что примерно 35 тыс. лет назад неандертальцы были вытеснены неоантропами - людьми современного физического склада, практически не отличающимися от сегодняшних европейцев, разве что имевшими более крепкое телосложение. Эти люди, получившие название кроманьонцев, населяли Европу, Африку, Китай. Их череп удивительно похож на череп современных людей и значительно отличается от черепа неандертальцев. С последними их роднят лишь массивный надглазный валик да крупные передние зубы. Кроманьонцы владели каменными орудиями (копьями, молотками, ножами) и были хорошо приспособлены к борьбе за существование. Считают, что они были непосредственными предками современного человека.
Дискуссионным вопросом является появление различных рас: то ли все они имеют общего предка (моноцентрическая гипотеза), то ли современные расы происходят от различных линий (полицентрическая гипотеза). Ясно одно: с появлением кроманьонцев, окончательно оформился человек, который за последние 35 - 40 тыс. лет практически не претерпел физиологической эволюции.














Вебинар для учителей

Свидетельство об участии БЕСПЛАТНО!
Полезное для учителя

Реализация образовательных программ осуществляется с применением исключительно электронного обучения и ДОТ

