
Россия, Калининград


СДЕЛАЙТЕ СВОИ УРОКИ ЕЩЁ ЭФФЕКТИВНЕЕ, А ЖИЗНЬ СВОБОДНЕЕ
Благодаря готовым учебным материалам для работы в классе и дистанционно
Скидки до 50 % на комплекты
только до
Готовые ключевые этапы урока всегда будут у вас под рукой
Организационный момент
Проверка знаний
Объяснение материала
Закрепление изученного
Итоги урока

Была в сети 06.06.2018 00:05

Мудрицкая Светлана Викторовна
заместитель директора, учитель биологии
76 лет
Местоположение
Специализация
Учебное пособие "Мир клетки" Модуль 4 "Основные принципы строения эукариотической клетки" к Программе "Путешествие в мир клетки"
Категория:
Биология
28.05.2018 21:30
Учебник:
Биология. Общая биология. Профильный уровень. 10 класс. Захаров В.Б. и др. М.: 2010. - 352 с.

Просмотр содержимого документа
«Учебное пособие "Мир клетки" Модуль 4 "Основные принципы строения эукариотической клетки" к Программе "Путешествие в мир клетки"»
2.2.3.7. Вакуоли растительных клеток 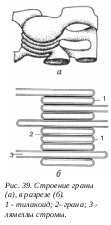 амелл стромы. Ламеллы стромы и мембраны тилакоидов образуются путем отделения от внутренней мембраны при начальных этапах развития пластид.
амелл стромы. Ламеллы стромы и мембраны тилакоидов образуются путем отделения от внутренней мембраны при начальных этапах развития пластид.
Клетки как низших, так и высших растительных организмов содержат в цитоплазме вакуоли, несущие ряд важных физиологических нагрузок. У молодых клеток может быть несколько мелких вакуолей, которые по мере роста и дифференцировки клетки сливаются друг с другом и образуют одну или несколько крупных вакуолей, занимающих до 90% объема всей клетки. Центральные вакуоли отделены от цитоплазмы одинарной мембраной, сходной по толщине с плазмалеммой. Мембрана, ограничивающая центральные вакуоли, носит название тонопласта. Возникают центральные вакуоли из мелких пузырьков, отщепившихся от аппарата Гольджи. Такие первичные вакуоли растут в объеме, сливаются друг с другом и образуют одну или несколько крупных вакуолей, оттесняющих цитоплазму с ядром и органоидами к периферии клетки. Полость вакуоли заполнена клеточным соком, представляющим собой водный раствор, в который входят различные неорганические соли, сахара, органические кислоты и их соли и другие низкомолекулярные соединения, а также некоторые высокомолекулярные вещества (например, белки).
Центральные вакуоли растений выполняют многообразные и важные функции. Одной из главных ее функций является поддержание тургорного давления клеток. Растворенные в соке вакуолей молекулы определяют его осмотическую концентрацию. Другая функция определяется тем, что вакуоль представляет собой большую полость, отделенную от гиалоплазмы мембраной, тонопластом, обладающим свойствами полупроницаемости и через который может происходить, как и через плазматическую мембрану, активный транспорт различных молекул. Поэтому вакуоли могут использоваться клетками как накопительные резервуары не только для отложения запасных веществ, но и для выброса метаболитов, для экскреции. В вакуолях происходит отложение многих глюкозидов, к которым относятся различные пигменты, например, антоцианы.
Другой обширный ряд функций вакуолей связан с накоплением запасных веществ, таких как сахара и белки. Сахара в вакуолях содержатся в виде растворов, встречаются и резервные полисахариды типа инулина. Запасание белков семян злаковых происходит в так называемых алейроновых вакуолях, которые заполняются альбуминами и глобулинами, после чего вакуоли обезвоживаются, превращаясь в твердые алейроновые зерна. При прорастании семян эти зерна обводняются и снова превращаются в вакуоли.
2.2.3.8. Сферосомы
Сферосомы - это мембранные пузырьки, встречающиеся в клетках растений. Они окрашиваются липофильными красителями, имеют высокий коэффициент преломления и поэтому хорошо видны в световой микроскоп. Сферосомы образуются из элементов эндоплазматической сети. На конце цистерны ЭПС начинает накапливаться осмиофильный материал, затем от этого участка отшнуровывается и начинает расти мелкий пузырек, достигающий диаметра 0,1-0,5 мкм. Рост сферосом и перестройка их содержимого связаны с накоплением в них масла, так что сферосома постепенно превращается в масляную каплю. Кроме жиров в составе сферосом обнаруживают белки и среди них фермент липазу, расщепляющую липиды.
2.2.3.9. Пероксисомы
Пероксисомы (микротельца) - это небольшие вакуоли (0,3-1,5 мкм), одетые одинарной мембраной, отграничивающей гранулярный матрикс, в центре которого располагается сердцевина. В зоне сердцевины часто, особенно в пероксисомах печеночных клеток, видны кристаллоподобные структуры, состоящие из регулярно упакованных фибрилл или трубочек. Изолированные сердцевины пероксисом содержат фермент уратоксидазу.
Пероксисомы обнаружены у простейших (амебы, тетрахимена), у низших грибов (дрожжи), у высших растений в некоторых эмбриональных тканях (эндосперм) и в зеленых частях, способных к фотореспирации, у высших позвоночных животных они обнаруживаются главным образом в печени и почках. В печени крыс на клетку число пероксисом колеблется от 70 до 100.
Пероксисомы часто локализуются вблизи мембран ЭПС. У зеленых растений пероксисомы не редко находятся в тесном контакте с митохондриями и пластидами.
Впервые пероксисомы были выделены из печени и почек. Во фракциях пероксисом обнаруживаются ферменты, связанные с метаболизмом перекиси водорода. Это ферменты (оксидазы, уратоксидаза, оксидаза d-аминокислот) окислительного дезаминирования аминокислот, при работе которых образуется перекись водорода (Н2О2) и каталаза, разрушающая ее. В пероксисомах печени каталаза составляет до 40 % всех белков и локализована в матриксе. Так как Н2О2 является токсическим веществом для клеток, то каталаза пероксисом может играть важную защитную роль. Пероксисомы цыплят и лягушек кроме уратоксидазы содержат ряд ферментов катаболизма пуринов.
У животных и некоторых растений (проростки клещевины) пероксисомы играют важную роль при превращении жиров в углеводы. В пероксидах происходит накопление специфических белков, которые синтезируются в цитозоле.
2.2.3.10. Митохондрии
Для осуществления любых клеточных функций необходима энергия. Клетки синтезируют молекулы АТФ (аденозинтрифосфат), обладающие высокоэнергетическими фосфатными связями. При разрушении этих связей выделяемая энергия может тратиться на любые клеточные функции: на активный транспорт веществ, на синтетические процессы, на механическую работу и т.д. В клетках животных синтез АТФ осуществляется специальными органеллами - митохондриями, в растительных клетках кроме митохондрий в энергообеспечении огромную роль играют хлоропласты. Митохондрии и пластиды - это двумембранные органеллы эукариотических клеток. Митохондрии присутствуют во всех к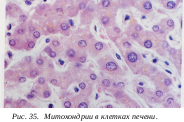 летках; их основная функция связана с окислением органических соединений и использованием освобождающейся при распаде этих соединений энергии в синтезе молекул АТФ. Поэтому митохондрии часто называют энергетическими станциями клетки.
летках; их основная функция связана с окислением органических соединений и использованием освобождающейся при распаде этих соединений энергии в синтезе молекул АТФ. Поэтому митохондрии часто называют энергетическими станциями клетки.
В
первые митохондрии были обнаружены Р. Альтманом, который описал эти клеточные органеллы в конце XIX века, называя их «биобластами». Митохондрии или хондриосомы (от греч. mitos– нить, chondrion- зернышко, soma- тельце) при световой микроскопии имеют вид гранул или нитей. Толщина митохондрий в различных клетках относительно постоянна (около 0,5 мкм), а длина колеблется, достигая у нитчатых форм до 7-60 мкм. В световом микроскопе на окрашенных препаратах не всегда можно определить реальный размер митохондрий.
Изучая митохондрии в электронном микроскопе, также трудно решить вопрос об их истинной длине, так как на срез попадает только незначительный объем митохондрии. Более того, на срезе одна извитая митохондрия может быть представлена несколькими сечениями, и только пространственная трехмерная реконструкция, построенная на изучении серийных срезов, может решить вопрос, имеем ли мы дело с тремя-шести отдельными митохондриями или же с одной изогнутой или разветвленной. Длинные ветвящиеся митохондрии были описаны в клетках культуры ткани млекопитающих, в клетках многих растений. Обычные же подсчеты показывают, что на печеночную клетку приходится около 200 митохондрий. Это составляет более 20% от общего объема цитоплазмы и около 30-35% от общего количества белка в клетке. П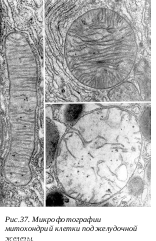 лощадь поверхности всех митохондрий печеночной клетки в 4-5 раз больше поверхности ее плазмолеммы. Локализация митохондрий в клетках быть различной. В малодифференцированных клетках меристемы растений митохондрии располагаются более или менее равномерно. В клетках эпителия почечных канальцев митохондрии ориентированы вдоль продольной оси клетки. Обычно митохондрии скапливаются вблизи тех участков цитоплазмы, где возникает потребность в АТФ. Так, в скелетных мышцах митохондрии находятся вблизи миофибрилл, в сперматозоидах образуют спиральный футляр вокруг оси жгутика. В аксонах нервных клеток митохондрии располагаются в синапсах, где происходит процесс передачи нервного импульса. Митохондрии независимо от их величины или формы имеют универсальное строение, их ультраструктура однообразна.
лощадь поверхности всех митохондрий печеночной клетки в 4-5 раз больше поверхности ее плазмолеммы. Локализация митохондрий в клетках быть различной. В малодифференцированных клетках меристемы растений митохондрии располагаются более или менее равномерно. В клетках эпителия почечных канальцев митохондрии ориентированы вдоль продольной оси клетки. Обычно митохондрии скапливаются вблизи тех участков цитоплазмы, где возникает потребность в АТФ. Так, в скелетных мышцах митохондрии находятся вблизи миофибрилл, в сперматозоидах образуют спиральный футляр вокруг оси жгутика. В аксонах нервных клеток митохондрии располагаются в синапсах, где происходит процесс передачи нервного импульса. Митохондрии независимо от их величины или формы имеют универсальное строение, их ультраструктура однообразна.
Митохондрии ограничены двумя мембранами. Наружняя митохондриальная мембрана отделяет ее от гиалоплазмы. Обычно она имеет ровные контуры, она представляет собой мембранный мешок. Наружнюю мембрану от внутренней отделяет межмембранное пространство шириной около 10-20 нм. Внутренняя мембрана ограничивает собственно внутреннее содержимое митохондрии, ее матрикс. Характерной чертой внутренней мембраны митохондрий является их способность образовывать многочисленные впячивания внутрь митохондрий. Такие впячивания чаще всего имеют вид плоских гребней, или крист. Матрикс митохондрий имеет тонкозернистое гомогенное строение, в нем иногда выявляются тонкие собранные в клубок нити (около 2-3 нм) и гранулы около 15-20нм. Теперь стало известно, что нити матрикса митохондрий представляют собой молекулы ДНК в составе митохондриального нуклеоида, а мелкие гранулы - митохондриальные рибосомы. Хондриом – это совокупность всех митохондрий в одной клетке.
2.2.3.11. Пластиды
Пластиды – это мембранные органоиды, встречающиеся у фотосинтезирующих эукариотических организмов (высшие растения, низшие водоросли, некоторые одноклеточные организмы). Подобно митохондриям, пластиды окружены двумя мембранами, в их матриксе имеется собственная геномная система, функции пластид связаны с энергообеспечением клетки, идущим на нужды фотосинтеза. У высших растений найден целый набор различных пластид (хлоропласт, лейкопласт, амилопласт, хромопласт), представляющих собой ряд взаимных превращений одного вида пластиды в другой. Основной структурой, которая осуществляет фотосинтетические процессы, является хлоропласт.
Хлоропласты – двумембранные органеллы дисковидной формы с шириной 2-4 мкм и протяженностью 5-10 мкм. У зеленых водорослей встречаются гигантские хлоропласты (хроматофоры), достигающие длины 50 мкм. Количество хлоропластов в клетках разных растений не одинаково. Так, у зеленых водорослей может быть по одному хлоропласту на клетку. Обычно на клетку высших растений приходится в среднем 10-30 хлоропластов. Встречаются клетки с большим количеством хлоропластов. Внешняя мембрана, как и внутренняя, имеет толщину около 7 нм, они отделены друг от друга межмембранным пространством около 20-30 нм. Внутренняя мембрана хлоропластов отделяет строму пластиды, аналогичную матриксу митохондрий. В строме зрелого хлоропласта высших растений видны два типа внутренних мембран. Это мембраны, образующие плоские, протяженные ламеллы стромы, и мембраны тилакоидов, плоских дисковидных вакуолей или мешков.
Л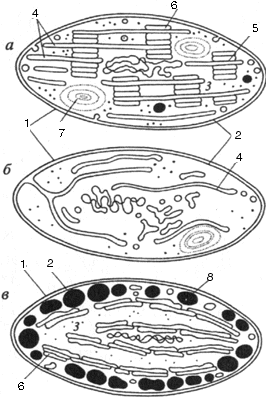 амеллы стромы (толщиной около 20 мкм) представляют собой плоские полые мешочки или имеют вид сети из разветвленных и связанных друг с другом каналов, располагающихся в одной плоскости. Обычно ламеллы стромы внутри хлоропласта лежат параллельно друг другу и не образуют связей между собой.
амеллы стромы (толщиной около 20 мкм) представляют собой плоские полые мешочки или имеют вид сети из разветвленных и связанных друг с другом каналов, располагающихся в одной плоскости. Обычно ламеллы стромы внутри хлоропласта лежат параллельно друг другу и не образуют связей между собой.
К
Рис. 38. Строение хлоропласта (а), лейкопласта (б), хромопласта (в)
1 – наружная мембрана; 2 – внут-ренняя мембрана; 3 – матрикс;
4 – лямеллы стромы; 5 – грана;
6 – тилакоид; 7 – крахмальное зерно; 8 – липидная капля с пигментами
роме мембран стромы в хлоропластах обнаруживаются мембранные тилакоиды. Это плоские замкнутые мембранные мешки, имеющие форму диска. Величина межмембранного пространства у них также около 20-30 нм. Такие тилакоиды образуют стопки наподобие столбика монет, называемые гранами. Число тилакоидов на одну грану варьирует: от нескольких штук до 50 и более. Размер таких стопок может достигать 0,5 мкм, поэтому граны видны в некоторых объектах в световом микроскопе. Количество гран в хлоропластах высших растений может достигать 40-60. Тилакоиды в гране сближены друг с другом так, что внешние слои их мембран тесно соединяются. Ламеллы стромы связывают между собой отдельные граны хлоропласта. Однако полости камер тилакоидов всезда замкнуты и не переходят в камеры межмембранного пространства ламелл стромы. Ламеллы стромы и мембраны тилакоидов образуются путем отделения от внутренней мембраны при начальных этапах развития пластид. В матриксе (строме) хлоропластов обнаруживаются молекулы ДНК, рибосомы; там же происходит первичное отложение запасного полисахарида крахмала в виде крахмальных зерен.
Хлоропласты - это структуры, в которых происходят фотосинтетические процессы, приводящие к связыванию углекислоты, синтезу сахаров и выделению кислорода.
Характерным для хлоропластов является наличие в них пигментов, хлорофиллов, которые и придают окраску зеленым растениям. При помощи хлорофилла зеленые растения поглощают энергию солнечного света и превращают ее в химическую. Поглощение света с определенной длиной волны приводит к изменению в структуре молекулы хлорофилла, она переходит при этом в возбужденное, активированное состояние. Освобождающаяся энергия активированного хлорофилла приводит к синтезу АТФ, энергия которой тратится на реакции синтеза сахаров.
Процесс развития различных пластид можно представить в виде идущего в одном направлении ряда смены форм:
Пропластида лейкопласт хлоропласт хромопластамилопласт.
Многими исследованиями был установлен необратимый характер онтогенетических переходов пластид. У высших растений возникновение и развитие хлоропластов происходят через изменения пропластид. Пропластиды представляют собой мелкие (0,4-1 мкм) двумембранные пузырьки, не имеющие отличительных черт их внутреннего строения. Они отличаются от вакуолей цитоплазмы более плотным содержимым и наличием двух отграничивающих мембран. Внутренняя мембрана может давать небольшие складки или образовывать мелкие вакуоли. Пропластиды чаще всего встречаются в делящихся тканях растений. Судьба таких пропластид будет зависеть от условий развития растений. При нормальном освещении пропластиды превращаются в хлоропласты. Сначала они растут, при этом происходит образование продольно расположенных мембранных складок от внутренней мембраны. Одни из них простираются по всей длине пластиды и формируют ламеллы стромы; другие образуют ламеллы тилакоидов, которые выстраиваются в виде стопки и образуют граны зрелых хлоропластов.
Лейкопласты отличаются от хлоропластов отсутствием развитой ламеллярной системы. Встречаются они в клетках запасающих тканей. Из-за их неопределенной морфологии лейкопласты трудно отличить от пропластид, а иногда и от митохондрий. Они, как и пропластиды, бедны ламеллами, но, тем не менее, способны к образованию под влиянием света нормальных тилакоидных структур и к приобретению зеленой окраски. В темноте лейкопласты могут накапливать различные запасные вещества, а в строме лейкопластов откладываются зерна вторичного крахмала. Другой формой пластид у высших растений является хромопласт, окрашивающийся обычно в желтый свет в результате накопления в нем каротиноидов. Хромопласты образуются из хлоропластов и значительно реже их лейкопластов (например, в корне моркови).
2.2.3.12. Клеточный центр и центриоли
Клеточный центр (центросомы) был обнаружен и описан более ста лет назад (Флемминг, 1875; Э. Бенеден, 1876). Клеточный центр характерен для клеток животных и низших растений. Отсутствует клеточный центр у высших растений, у низших грибов и некоторых простейших. Обычно эта органелла располагается в геометрическом центре клетки, поэтому и названа клеточным центром. Наиболее типичное строение клеточный центр имеет в клетках животных: он представляет собой зону, состоящую из центриолей, окруженных фибриллярной массой.
Тонкое строение центриолей удалось изучить с помощью электронного микроскопа. Основу строения центриолей составляют расположенные по окружности девять триплетов микротрубочек, образующие полый цилиндр (шириной около 0,15 мкм, а длиной - 0,3-0,5 мкм). Каждый триплет микротрубочек располагается к радиусу цилиндра под углом около 400.
С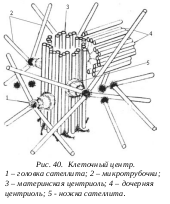 истему микротрубочек центриоли обычно описывают формулой 9 + 0, или (9х3) + 0, подчеркивая отсутствие микротрубочек в её центральной части.
истему микротрубочек центриоли обычно описывают формулой 9 + 0, или (9х3) + 0, подчеркивая отсутствие микротрубочек в её центральной части.
Вокруг каждой центриоли расположен бесструктурный, или тонковолокнистый матрикс. Около центриолей можно обнаружить несколько дополнительных структур: это сателлиты, фокусы схождения микротрубочек, исчерченные волокнистые корешки и дополнительные микротрубочки, образующие вокруг центриоли особую зону - центросферу.
Обычно в интерфазных клетках присутствуют две центриоли, располагающиеся рядом друг с другом, образуя дуплет центриолей, или диплосому. В диплосоме центриоли располагаются под прямым углом по отношению друг к другу. Из двух центриолей различают «материнскую» и «дочернюю». В дистальном участке материнской центриоли располагается аморфный материал в виде выростов или шпор - это придатки. Их нет на дочерней центриоли. Только материнская центриоль содержит сателлиты, состоящие из конусовидной ножки, расположенной на стенке центриоли, и головки, заканчивающейся на этой ножке. Ножки сателлитов часто имеют поперечную исчерченность. Микротрубочки отходят и от головок сателлитов. Эти центросомные микротрубочки не отходят непосредственно от микротрубочек цилиндров центриолей, а связаны или с сателлитами, или с матриксом. Такие микротрубочки и образуют лучистую сферу - центросферу вокруг центриоли.
Центриоли в делящихся клетках принимают участие в формировании веретена деления и располагаются на полюсах клетки. В неделящихся клетках центросомы часть определяют полярность клеток эпителия и располагаются вблизи аппарата Гольджи.
2.2.4. Ядро эукариотических клетокСам термин «ядро» впервые был применен М. Брауном в 1833 г. для обозначения шаровидных постоянных структур в клетках растений. Позднее такую же структуру описали во всех клетках высших организмов. Большинство клеток содержат одно ядро, хотя описаны и многоядерные клетки. Эритроциты млекопитающих и зрелые ситовидные клетки растительных организмов не имеют ядра; часто в популярной литературе безъядерными клетками называют тромбоциты, которые являются не клетками, а фрагментами мегакариоцита.
Клетки печени, поджелудочной железы, нервные клетки имеют ядра шаровидной формы, у миоцитов ядра дисковидные. Клетки гранулоцитов крови имеют сегментированные ядра.И тем не менее, самые непохожие по форме ядра, имеют общий план строения. Клеточное ядро состоит из ядерной оболочки, отделяющей его от цитоплазмы, хроматина, ядрышка, ядерного белкового остова (матрикса) и нуклеоплазмы (или ядерного сока). Эти основные компоненты встречаются практически во всех неделящихся клетках эукариотических одно- или многоклеточных организмов.
Ядро выполняет функцию хранения генетической информации, а также обеспечивает реализацию этой генетической информации через управление синтезом белка.
54








© 2018, Мудрицкая Светлана Викторовна 453 9
Рекомендуем курсы ПК и ППК для учителей






Похожие файлы
Вебинар для учителей

Свидетельство об участии БЕСПЛАТНО!
Полезное для учителя

Реализация образовательных программ осуществляется с применением исключительно электронного обучения и ДОТ

