
Россия, Моздок


СДЕЛАЙТЕ СВОИ УРОКИ ЕЩЁ ЭФФЕКТИВНЕЕ, А ЖИЗНЬ СВОБОДНЕЕ
Благодаря готовым учебным материалам для работы в классе и дистанционно
Скидки до 50 % на комплекты
только до
Готовые ключевые этапы урока всегда будут у вас под рукой
Организационный момент
Проверка знаний
Объяснение материала
Закрепление изученного
Итоги урока

Был в сети 19.04.2024 23:14

Штарк Константин Анатольевич
методист по естественнонаучному направлению
47 лет
Местоположение
Специализация
Исследовательский проект «Растительные жгутиковые: филогенез фоторецепции и подвижности»
Категория:
Биология
01.05.2019 18:14

Просмотр содержимого документа
«Исследовательский проект «Растительные жгутиковые: филогенез фоторецепции и подвижности»»
15
X Региональный конкурс молодых исследователей "Ступень в науку".
Секция: Общая биология и биоэкология
Тема: «Растительные жгутиковые: филогенез фоторецепции и подвижности»
Авторы работы: Домский Никита Анатольевич, Мкртчян Мария Кареновна
Место выполнения работы:
МБОУ СОШ №108, 10 класс, МБОУ СОШ №3, 9 класс, СЮН, г. Моздок
Научный руководитель:
Штарк Константин Анатольевич,
педагог Станции юных натуралистов Моздокского района
Моздок, 2012-2013
Оглавление.
| I | Введение | 3 |
| II | Основная часть | 5 |
| III | Заключение | 11 |
| IV | Библиографический список | 13 |
| V | Приложение | 14 |
Введение.
Принимая во внимание уже известное из изучаемой нами проблематики, а также наши предположения мы сформулируем следующую цель нашего исследования: выявить возможные взаимосвязи в развитии светочувствительного глазка и органоидов активного движения клетки. В соответствии с данной целью можно поставить следующие задачи:
а) показать, что подвижность в природе развивается благодаря развитию органоидов или органов рецепции, а не с развитием нервной системы, как считалось ранее;
б) предложить и обосновать понятие юнирефлекса, как явления причинно-следственной связи между рецептирующими структурами одноклеточных организмов и органоидом активного движения (рабочим органоидом);
в) поставить опыт, позволяющий с большой долей вероятности предположить, какая поляризация развивается в одноклеточном в результате его реакции на свет.
Для выполнения исследования мы использовали метод эксперимента и наблюдения за водорослями в световой микроскоп. Для этого нам необходимы следующие материалы и оборудование:
5 проб воды из различных (в том числе искусственных) водоёмов, предположительно содержащие одноклеточные зелёные водоросли, пипетка, предметное стекло, покровное стекло, микроскоп с длиной тубуса 250 мм, объектив с увеличением 40 единиц, окуляр с увеличением 20 единиц- общее увеличение- 800 единиц. Из каждой пробы воды мы готовим, поочерёдно, 10 препаратов. Данные по встречаемости жгутиконосцев в каждом препарате собраны в таблице №1.
Согласно данным справочника по физике (2) освещённость выражается формулой:
E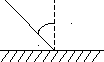 =J cos /l2
=J cos /l2
J – сила света
- угол падения
l – расстояние до источника света
Единицы освещенности:
Люкс (лк) и фот
1 фот=104 лк.
Освещённость вблизи окна в яркий солнечный день- 1000 лк, в пасмурный- 100 лк.
Для выполнения работы мы три раза посещали лабораторию в санэпидемстанции. В первый день, 13 сентября, погода была облачная, и к началу работы, в 1500 освещенность по нашим примерным оценкам составила не более 200 лк. Мы рассмотрели по два препарата из каждой пробы. Все из наблюдаемых 14 жгутиконосцев постоянно передвигались, более или менее активно.
Во второй день наблюдения, 1 октября, был солнечный день, и в 1300 освещенность вблизи окна лаборатории составляла, примерно, 1000 лк. Мы рассмотрели по пять препаратов из каждой пробы (40 жгутиконосцев исследуемых родов). Большинство из них перемещались заметно медленнее, чем в первый день эксперимента, многие иногда замедляли движение до полной остановки.
В третий день, 4 октября, наблюдения день был пасмурный, облачный, и в 1300 освещенность в лаборатории составляла, примерно, 100-150 лк. Мы рассмотрели по три препарата из каждой пробы, в которых удалось наблюдать ещё 29 жгутиконосцев. Одноклеточные снова вели себя активно, как будто "искали" свет.
Основная часть.
Что же в процессе эволюции появляется раньше- рецепторы или нервная система? Следуя биогенетическому закону Мюллера и Геккеля, говорящего о том, что онтогенез есть краткое повторение филогенеза, приведем косвенное подтверждение того, что рецепция появилась раньше, чем нервная деятельность.
Некоторые исследователи (15) считают, что один из самых первых и самых выраженных признаков возникновения оси тела у позвоночных – это появление на спинной поверхности зародыша утолщения эктодермы – нервной пластинки, образование которой индуцируется подлежащей мезодермой. Глаз состоит главным образом из нервной ткани и поэтому развивается целиком из эктодермы; мезодерма же участвует лишь в формировании наружных защитных оболочек и системы кровоснабжения. (10,12,16)
Следуя вышеизложенному, логично предположить, что, поскольку эктодерма первична в происхождении по отношению к мезодерме, сетчатка глаза, развиваясь из эктодермы, при реципрокном взаимодействии, в первую очередь с эктодермой, является более самостоятельным образованием, зависимым в основном от характера взаимодействия в первом зародышевом листке, чем нервная пластинка. Постараемся теперь философски обосновать первичность рецепции по отношению к нервной деятельности. Известно, что на возникновение и развитие нервных структур организм затрачивает энергии больше, чем на существование каких-либо других его структур. Опираясь на учение даосов, равновесно-интегративную теорию эволюции (Спенсер, Ле Дантек, Уорд) (5), взгляды А.А. Кропоткина (9) и В.С. Соловьева, В.И. Вернадского и др. можно говорить о первичности содействий по отношению к противодействиям в природе и об эволюции в ней по схеме:
равновесие 1 – неравновесие – равновесие 2.
Исходя из законов термодинамики, равновесие системы – это её состояние, когда А=А'=0. В такой или близкой к такой системе энергия сохраняется благодаря высокому уровню её обмена с внешней средой (много отдает – много получает). В нервной же системе, главное значение которой согласование работы по жизнеобеспечению организма, энергия в форме солитонной волны возбуждения сохраняется, благодаря поглощению наибольшего, в сопоставлении с другими системами организма, количества энергии, образующейся в результате энергетического обмена.
Другими словами, количество энергии, потребляемое нервной системой гораздо больше количества энергии ею отдаваемой (тем более это относится к нервной системе позвоночных, т.к. распространение импульса в ней происходит без декремента, благодаря миелинизации её проводящих отростков). Следовательно, нервная система характеризуется наиболее неравновесным состоянием по отношению к другим системам и не могла, согласно указанной выше формуле эволюции, появиться ранее рецептирующих структур. (11) Более того, поскольку нервная клетка получает импульс от рецепторной, отдающей его (т.е. энергию), а не передающей (как нервная клетка), вероятно, что именно появление нервной системы является прямым следствием развития рецептирующих клеток. Действительно, молекулярные механизмы нервной системы – ионные каналы, медиаторы и их рецепторы, - как сейчас считают, были сформированы на более ранних (возможно, на "донервных") стадиях филогенеза, так как они практически не различаются у животных разного уровня развития. (11) Следовательно, подвижность как свойство живого вещества связано с возникновением и развитием чувствительных структур, а не нервной системы. Характерно, что многие организмы с диффузной системой или типа ортогон не совершают активных движений.
В соответствии с вышеизложенным возникает вопрос: "Как же связаны фоторецепция и активная подвижность в своей коэволюции, если таковая действительно существует?"
Известно, что суточные и годовые ритмы организмов основываются на системе свободного отсчета времени по принципу биологических часов. В природных условиях эта система находится под контролем внешних факторов-синхронизаторов, среди которых у различных групп организмов главная роль принадлежит фотопериоду. (3,4,6)
Фотопериодический контроль нервно-секреторной активности гипоталамуса в естественных условиях происходит на базе зрительной регуляции. Однако даже это необязательное условие экспериментально показало, что изменение режима освещения могут восприниматься и без участия органов зрения на базе светочувствительных структур центральной нервной системы. В частности в опытах с американскими древесными воробьями Spizella arborea показано, что фотостимуляция глаз возможна и у животных хирургически лишенных органов зрения и эпифиза (F. Wilson. 1991 г.). Подобные опыты были и ранее проведены на других видах птиц, а также на амфибиях и ящерицах. В научном мире активно исследуются причинно- следственные связи фотопериода с генотипом организма. (7)
Еще в конце 40-х годов нашего столетия профессор Московского университета Д.А. Сабинин высказал предположение о связи механизма суточной физиологической периодики со структурой генетического аппарата. В конце 60-х годов такая гипотеза была возрождена в виде концепции хронона (С. Еhret, Е. Тrucco, 1967г.) (18).
Согласно этой концепции материальным носителем счёта времени служит длинная молекула ДНК, нити которой расходятся и на них строятся информационная (матричная) РНК, достигающая полной длины одиночной нити ДНК примерно за 24 часа. Процесс сопровождается рядом сопутствующих реакций; он не зависит от температуры в очень широком диапазоне её колебания. (7)
Исходя из сказанного, "первичная" рецепция происходит не на клеточном, а на молекулярно-генетическом уровне организации живого. Именно этот уровень организации обеспечивает надежность функционирования рефлекторных дуг. Если рецептор и рабочий орган по каким-либо причинам "выйдут из строя", то рефлекторная дуга останется в виде ее трёх центральных звеньев. Данное обстоятельство показывает, что рецепторы могут существовать без рабочего органа, т.е. рецепция может существовать без движения; появляется ранее движения и связано с ним причинно-следственными отношениями (взаимодействие в рефлекторной дуге не является непричинными формами отношения, т.к. все её части имеют общее происхождение, причём, как правило, все части, кроме рабочего органа, происходят из первого зародышевого листка, т.е. является причиной возникновения рабочего органа).
Известно также, что рецептирующие структуры функционально подвижнее структур "рабочих", физиологическая деятельность которых порождает активные движения организма (внутренние или внешние) (8). Это обстоятельство показывает, что рецептирующие структуры являются более внутренним качеством по отношению к двигательным структурам. Согласно идее о содействии, как движущей силе прогресса в едином качестве достаточно появления малых противодействий, малого нарушения равновесия (выражается в разности ПП и ПД), чтобы возникло возбуждение. Последнее вредно для клетки, поэтому волна возбуждения по градиенту количества вещества распространяется к жгутику жгутиконосца, где преобразуется в активное движение. Вероятно, для минимизации декремента волны импульса возбуждения и фоторецептор расположен на узелке жгутика, т.е. дуга рецептор – нуклеиновые кислоты – рабочий органоид является почти замкнутой.
Итак, опираясь на описанные ранее физиологические механизмы взаимодействия рецептирующих структур и органоидов движения, можно предложить вести речь о рефлекторной деятельности на клеточном уровне организации. Для удобства мы предлагаем ввести термин юнирефлекс, по аналогии с предложенным П. Карсом термином – юнимышление. (13). У жгутиконосцев, исследуемых нами, очевидна рефлекторная, или юнирефлекторная дуга, состоящая из сложных рецептирующих структур, солитонной волны (1), распространяющейся по возбудимым проводящим стркутурам клетки к ядру; здесь, нуклеиновыми кислотами, производится анализ полученного сигнала и синтез необходимого ответа (именно с квантовой солитонно-голографической природой ДНК и РНК. П. Карс и другие (Гаряев, Тертышный и т.д.) связывают индукцию и осуществление основных процессов юнимышления), затем, по проводящим путям синтезированный сигнал поступает в рабочий органоид.
Подобный взгляд не противоречит выделенным И.П. Павловым трём основным этапам рефлекторной деятельности.
Во-первых, юнирефлекс клетки причинно обусловлен действием раздражителя. Жгутиковые, обладая положительным и отрицательным таксисами ведут себя целесообразно, причем это поведение характеризуется отношением временной асимметрии и единой квантовой солитонно-голографической природой волн возбуждения, раздражителя и структур юнирефлекторной дуги, а, значит, является следствием действия раздражителя.
Во-вторых, поступающая в клетку жгутикового информация оценивается, сопоставляется, т.е. анализируется, а затем синтезируется. Имея фототаксис и хемотаксис, всегда предпочтение отдается фототаксису и автотрофному, обеспечивающему наибольшим количеством энергии, способу питания.
В-третьих, соблюдается принцип структурности, т.к. все физиологические процессы юнирефлекса протекают в соответствующих морфологических структурах клетки.
Итак, результаты многократно повторяемых экспериментов показывают, что клетки жгутиковых активны в темноте, а на свету замедляются, успокаиваются. Сходные данные описаны Шмидт-Ниельсоном (14). Известно, что у позвоночных животных возбудителем фоторецепторов является также темнота, на их клеточных мембранах, несколько деполяризованных в темноте, в ответ на освещение возникает гиперполяризация, а у беспозвоночных – деполяризация. Сходная с позвоночными реакция на свет у жгутиковых водорослей наводит на мысль, что и на мембранах последних развивается гиперполяризация. Это обстоятельство показывает, что биогенетический закон выполняется не только морфологически: в эволюции циллиарного типа организации фоторецепторов, но и физиологически.
Естественно, что гиперполяризация ведет к успокоению клетки, т.е.к "суперпокою". Волна возбуждения гаснет, что обеспечивает максимальное усвоение клеткой солнечной энергии. Вероятно, что в темноте клетки жгутиковых деполяризованы и волны возбуждения преобразуются в активные движения жгутиковых (поскольку выделение медиаторов, по аналогии с многоклеточными организмами нецелесообразно).
Вероятно, что гиперполяризация или просто потенциал покоя жгутиконосцев невозможна в темноте, потому, что тогда фототаксис этих одноклеточных был бы отрицательным, и они не имели бы хлоропластов и не были бы автотрофными организмами. Данные выводы подтверждают тезис о том, что любая термодинамическая система стремится к равновесию, т.е. состоянию покоя. Гетеротрофность развилась, вероятно, как следствие раздражения темнотой, как способ частичного возмещения энергии, затраченной на активное движение в темноте.
Заключение.
1. Светочувствительный глазок – органоид "зрения" впервые появляется у одноклеточных монадных водорослей, т.е. у автотрофных организмов.
2. Благодаря этому у них эволюционирует органоид активного движения – жгутик, т.к. возбуждение их родопсина и импульсы (волны) возникающие в результате переполяризации их внутренней среды, реализуют свою энергию в реактивности, в первую очередь, возбудимых структур клетки, таких как жгутик. Одноклеточные водоросли не имеющие фоторецептирующих структур, не имеют и органоидов активного движения. Клетки со светочувствительным глазком изначально имели жгутик.
3. Следовательно, движение как свойство живого вещества связано с возникновением и развитием чувствительных структур, а не нервной системы. Характерно, что многие организмы с диффузной системой или типа ортогон не совершают активных движений.
4. Вероятно, нервная система связана с появлением гетеротрофности, т.к. в данных организмах высока степень деградации энергии и для сохранения координации и уровня энергоинформационного обмена возникла нервная система.
5. Благодаря фоторецепции и органоиду зрения, жгутиконосцы функционально подвижнее клеток многоклеточных растений (разность между ПП и ПД у жгутиконосцев меньше).
6. Ответную реакцию одноклеточного организма на раздражение, осуществляемую и контролируемую посредством солитонного юнимышления клетки (П. Карс, 1998) мы предлагаем назвать по аналогии юнирефлексом, т.к. налицо дуга, состоящая из рецепторного органоида, солитонной волны возбуждения контролирующей и обеспечивающей надежность любого рефлекса молекулы ДНК и рабочего органоида (органоидов). Именно молекулы ДНК и РНК, синтезированные за 24 часа являются первым звеном реакции организма на фотопериод (Ehret, Trucco, 1967).
7. Одноклеточная водоросль активна в темноте, где на её плазматической мембране вероятно развивается деполяризация, На свету их клетки успокаиваются, вероятно, на мембране в результате фототрансдукции развивается гиперполяризация.
Библиографический список.
А.Т. Филиппов. Многоликий солитон. М., "Наука", 1990.
А.С. Енохович. Справочник по физике и технике. М.,Просвещение, 1989.
В.В. Полевой. Физиология растений. М. "Высшая школа", 1989,
В.И. Артамонов. Занимательная физиология растений. М., 1991.
Г. Спенсер. "Гипотеза развития" // "Теория развития". СПб., 1904.
Жизнь растений, т.3, под ред.проф. М.М. Голлербаха. М.Просвещение, 1977.
И.А. Шилов. Экология. М., Высшая школа, 1998.
Коган А.Б., Косицкий Г.И. и др. Физиология человека и животных.т.2, М., Высшая школа, 1984.
Кропоткин П.А. "Взаимная помощь среди людей и животных как двигатель прогресса", Пг, 1992.
М.А. Островский М.А., Молекулярная физиология зрения: системы фоторецепции и защиты от фотоповреждения // Природа.М., 1993.
Ноздрачев А.Д. Общий курс физиологии человека и животных, т.1. М. Высшая школа, 1995.
Островский М.А., Говардовский В.И. Механизмы фоторецепции позвоночных. // Физиология зрения. М., 1992.
П. Карс // Природа и человек, 1998, №3.
Шмидт-Ниельсон. Сравнительная физиология организмов. В 3-х т.
Э. Дьюкар. Клеточные взаимодействия в развитии животных. Под ред. доктора биол. наук Т.А. Детлаф. М., 1978.
Dowling J. The Retina. Cambridge – London, 1987
Coulombre A.J., 1965. The eye In: Organogenesis (DeHaah R.L. and Ursprung H., eds.), pp. 219-251, Holt, Rinehart and Winston, New York, Toronto, London.
Ehret C. F., Trucco E. Molecular models for the Circadian Clock. J. The Chronon concept. – J. Theoret. Biology, 1967, p. 15.
Приложение.
Таблица №1
| № пробы | Место, где взята проба | № препарата | Количество жгутиковых в препарате (подсчет на свету) | Род исследуемых жгутиковых |
| 1 | 2 | 3 | 4 | 5 |
| 1. | Аквариум в здании СЮН | 1 | 0 |
|
| 2 | 0 |
| ||
| 3 | 0 |
| ||
| 4 | 0 |
| ||
| 5 | 1 | хламидомонада | ||
| 6 | 0 |
| ||
| 7 | 2 | хламидомонада | ||
| 8 | 1 | хламидомонада | ||
| 9 | 1 | хламидомонада | ||
| 10 | 0 |
| ||
| 2. | Водная станция | 1 | 0 |
|
| 2 | 0 |
| ||
| 3 | 0 |
| ||
| 4 | 0 |
| ||
| 5 | 0 |
| ||
| 6 | 0 |
| ||
| 7 | 0 |
| ||
| 8 | 0 |
| ||
| 9 | 0 |
| ||
| 10 | 0 |
| ||
| 3. | Озеро на УОУ СЮН | 1 | 0 |
|
| 2 | 2 | хламидомонада | ||
| 3 | 2 | эвглена | ||
| 4 | 1 |
| ||
| 5 | 1 | хламидомонада | ||
| 6 | 1 | эвглена, хламидомонада | ||
| 7 | 1 | эвглена | ||
| 8 | 3 | эвглена | ||
| 9 | 2 | эвглена, хламидомонада | ||
| 10 | 5 | эвглена, хламидомонада | ||
| 4. | Домашний аквариум | 1 | 0 |
|
| 2 | 1 | хламидомонада | ||
| 3 | 2 | эвглена, хламидомонада | ||
| 4 | 0 |
| ||
| 5 | 1 | эвглена | ||
| 6 | 2 | хламидомонада | ||
| 7 | 3 | эвглена, хламидомонада | ||
| 8 | 1 | хламидомонада | ||
| 9 | 2 | эвглена | ||
| 10 | 0 |
| ||
| 5. | Дренажный канал в Алборовском лесу
| 1 | 7 | эвглена |
| 2 | 4 | эвглена | ||
| 3 | 6 | эвглена, хламидомонада | ||
| 4 | 3 | эвглена | ||
| 5 | 6 | эвглена, хламидомонада | ||
| 6 | 2 | эвглена | ||
| 7 | 6 | эвглена, хламидомонада | ||
| 8 | 5 | эвглена | ||
| 9 | 5 | эвглена | ||
| 10 | 7 | эвглена, хламидомонада | ||
|
| Итого | 50 | 83 |
|














Вебинар для учителей

Свидетельство об участии БЕСПЛАТНО!
Полезное для учителя

Реализация образовательных программ осуществляется с применением исключительно электронного обучения и ДОТ

