
Россия, Калининград


СДЕЛАЙТЕ СВОИ УРОКИ ЕЩЁ ЭФФЕКТИВНЕЕ, А ЖИЗНЬ СВОБОДНЕЕ
Благодаря готовым учебным материалам для работы в классе и дистанционно
Скидки до 50 % на комплекты
только до
Готовые ключевые этапы урока всегда будут у вас под рукой
Организационный момент
Проверка знаний
Объяснение материала
Закрепление изученного
Итоги урока

Была в сети 17.01.2021 23:39

Сафронова Марина Владимировна
преподаватель биологии, химии и экологии
35 лет
Местоположение
Специализация
Структура и динамика популяций (демэкология)
Категория:
Экология
09.04.2019 14:24

Просмотр содержимого документа
«Структура и динамика популяций (демэкология)»
ТЕМА: Структура и динамика популяций (демэкология)
ПЛАН:
1. Понятие о популяции в экологии
2. Основные характеристики популяции.
3. Структура популяций
3.1.Пространственная и этологическая структуры популяций
3.2. Половая и возрастная структура популяции
4. Динамика популяций
4.1. Кривые выживания
4.2. Рост популяции и кривые роста
4.3. Колебания численности популяции
1. Понятие о популяции в экологии
Популяцией в экологии называют группу особей одного вида, находящихся во взаимодействии между собой и совместно населяющих общую территорию. Слово “популяция” от лат. populus – народ, население. Экологическую популяцию, определяют как население одного вида на определенной территории.
Члены одной популяции оказывают друг на друга не меньше воздействие, чем физические факторы среды или другие, совместно обитающие, виды организмов. В популяциях проявляются в той или иной степени все формы связи, характерные для межвидовых отношений, но наиболее ярко выражены мутуалистические (взаимно полезные) и конкурентные. Специфические внутривидовые взаимосвязи – это отношения, связанные с воспроизводством между особями разных полов и между родителями и дочерним поколениями.
В популяциях действуют законы, позволяющие использовать ограниченные ресурсы среды для обеспечения воспроизводства потомства. Достигается это в основном через количественные изменения населения.
Поддержание оптимальной, в данных условиях, численности называют гомеостазом популяции. У каждой популяции можно выделить основные системные атрибуты (характеристики): состав, структуру и функцию.
Таким образом, популяции, как групповые объединения, обладают рядом специфических свойств. Групповые особенности – это основные характеристики популяций. К ним можно отнести:
- численность популяции;
- плотность популяции;
- рождаемость;
- смертность популяции.
2. Основные характеристики популяции
Численность популяции – это общее количество особей на данной территории или в данном объеме. Оно никогда не бывает постоянно и зависит от соотношения интенсивности размножения (плодовитости) и смертности. В процессе размножения происходит рост популяции, смертность же приводит к сокращению ее численности.
Плотность популяции – определяется количеством особей (либо биомассой) на единице площади или объема, занимаемого популяцией.
Существует много методов определения численности и плотности популяции, но все их можно объединить в 4 основные группы: метод прямого подсчета, мечения и повторных отловов, выборные и косвенные методы.
Рождаемость – (плодовитость) определяется числом новых особей, появившихся за единицу времени в результате размножения. Низкая плодовитость характерна для тех видов, которые проявляют большую заботу о потомстве. Кроме того, плодовитость зависит от скорости созревания, числа генераций в год, соотношения в популяции самцов и самок, обеспеченности кормом, влияния погодных условии и др. факторов.
Смертность популяции – это количество особей, погибших за определенный период. Различают три типа смертности. Первый характеризуется одинаковостью во всех возрастах; второй- повышенной гибелью особей на ранних стадиях развития; третий тип характеризуется повышенной гибелью взрослых (старых) особей.
Факторы смертности разнообразные. Это в основном: физические условия (низкие и высокие температуры, ливневые осадки, засуха и др.), биологические факторы (отсутствие корма, заболевания, и др.) и антропогенные (загрязнение окружающей среды, вырубка лесов, охота и др.).
3. Структура популяций
Ввиду сложной организации, различают: половую, возрастную, пространственную и этологическую структуру популяций.
3.1.Пространственная и этологическая структуры популяций
Пространственная структура. Каждая популяция занимает пространство, обеспечивающее условия жизни для ограниченного числа особей. При изучении пространственной структуры различают случайное, равномерное и неравномерное (групповое) распределения особей на территории (в пространстве).
Случайное распределение в природе встречается редко; оно наблюдается в случаях, когда среда очень однородна, а организмы не стремятся объединиться в группы. Равномерное распределение бывает там, где между особями очень сильна конкуренция или существует антагонизм. Наиболее часто наблюдается неравномерное (групповое) распределение — образование различных скоплений.
Активность особей, пар и семейных групп у позвоночных и высших беспозвоночных обычно ограничена определенной зоной, называемой индивидуальным (или семейным) участком территории. У высших животных внутрипопуляционное распределение регулируется системой инстинктов. Им свойственно особое территориальное поведение — реакция на местонахождение других членов популяции.
В зависимости от характера использования пространства подвижных животных подразделяют на оседлых и кочевых. Оседлые животные в течение всей или большей части жизни используют довольно ограниченный участок среды. Им присущи инстинкты привязанности к своему участку, регулярное возвращение к месту размножения после длительных и дальних миграций.
Кочевые животные совершают постоянные передвижения в пространстве, так как они зависят от запаса корма на данной территории. Кочевой образ жизни характерен преимущественно для стад и стай (кочующие группы многих рыб во время нагульных миграций, стада слонов, зебр, антилоп, северных оленей и т. п.). Перемещение кочующих видов по площади обычно происходит за сроки, достаточные для восстановления кормовых ресурсов на пастбищных участках. Масштабы и длительность таких миграций определяют обилие пищи и численность стада.
Этологическая (поведенческая) структура. Она отражает разнообразные формы совместного существования особей в популяциях. Одиночный образ жизни следует выделить в первую очередь, хотя полностью одиночного существования организмов в природе нет, так как в этом случае было бы невозможно размножение. Семейный образ жизни — усиливаются связи между родителями и потомством, начинает заметно проявляться территориальное поведение животных. Путем различных сигналов, маркировки, угроз и тому подобного обеспечивается владение участком, достаточным для выкармливания потомства.
Стая — временное объединение животных, проявляющих биологически полезную организованность действий (для защиты от врагов, добычи пищи, миграции и т. п.). Наиболее широко стайность распространена среди рыб и птиц, хотя встречается и у млекопитающих (например, у собак).
Стадо — длительное или постоянное объединение животных, в котором осуществляются все основные функции жизни вида: добывание корма, защита от хищников, миграция, размножение, воспитание молодняка.
Основу группового поведения в стадах составляют взаимоотношения доминирования. Характерно наличие временного или относительно постоянного лидера, которому подражают особи стада.
Активное руководство стадом (специальные сигналы или угрозы) осуществляют вожаки. Иерархически организованному стаду свойственен закономерный порядок перемещения, определенные позиции при защите от врагов, расположение на местах отдыха и др. (рис. 4.3).
Колония — это групповое поселение оседлых животных на длительное время или на период размножения. По сложности взаимоотношений между особями колонии очень разнообразны, наиболее сложные отношения складываются в поселениях для общественных насекомых (термитов, муравьев, пчел, ос и др.), возникающие на основе сильно разросшейся семьи. Члены колоний постоянно обмениваются информацией друг с другом.
3.2. Половая и возрастная структура популяции
Средняя продолжительность жизни организмов и соотношение численности (или биомассы) особей различного возраста характеризуются возрастной, а соотношение особей разного пола — половой структурами популяции. Соотношение особей по полу и особенно доля размножающихся самок в популяции имеют большое значение для дальнейшего роста ее численности. Соотношение полов в популяции устанавливается по законам генетики, по поведенческим различиям полов и, в известной мере, под влиянием среды.
Формирование возрастной структуры происходит в результате совместного действия процессов размножения и смертности.
Со временем даже в пределах одной и той же популяции могут происходить значительные изменения возрастной структуры. В таких случаях «включаются механизмы», автоматически возвращающие популяцию к некоторому нормальному для данной популяции возрастному распределению. Причем популяции, включающие в себя много возрастных групп, в меньшей степени подвержены воздействию факторов, влияющих на размножение в конкретном году.
Даже крайне неблагоприятные условия, вызывающие полную гибель приплода в тот или иной год, не становятся катастрофическими для популяции сложной структуры, так как родительские пары участвуют в воспроизводстве многократно. Тем не менее следы изменений условий жизни сохраняются в облике сложных популяций гораздо дольше, чем у простых. Для роста численности популяций большое значение имеет соотношение особей по полу. Генетический механизм обеспечивает примерно равное соотношение особей разного пола при рождении. Однако исходное соотношение вскоре нарушается в результате различий физиологических, поведенческих и экологических реакций самцов и самок, вызывающих неравномерную смертность.
Анализ возрастной и половой структуры популяции позволяет прогнозировать ее численность на ряд ближайших поколений и лет. Этим пользуются для оценки возможности промысла рыбы, в охотничьих хозяйствах и прочих подобных случаях.
С возрастом требования особи к среде и устойчивость к отдельным ее факторам закономерны и весьма существенно изменяются. Возрастная структура отражает соотношение различных возрастных групп в популяции.
По Г. А. Новикову выделяется пять возрастных групп животных:
1. новорожденные – до момента прозрения;
2. молодые – подрастающие особи, еще не достигшие половой зрелости;
3. полувзрослые – близкие к половой зрелости;
4. взрослые – половозрелые животные;
5. старые – переставшие размножаться особи.
Обычно в популяции присутствуют все возрастные группы, между ними наблюдаются постепенные переходы.
Возрастная структура популяций имеет приспособительный характер.
Таким образом, популяции свойственна определенная организация.
Распределение особей по территории, соотношение групп по полу, возрасту, морфологическим, физиологическим, поведенческим и генетическим особенностям, отражают структуру популяции. Она формируется, с одной стороны, на основе общих биологических свойств вида, а с другой – под влиянием абиотических факторов среды и популяций других видов. Структура популяций имеет, следовательно, приспособительный характер. Кроме адаптивных возможностей она характеризуется еще и приспособительными чертами групповой организации.
Приспособления организмов в конечном итоге направлены на повышение вероятности выжить и оставить потомство. Это мы называем экологической стратегией.
Все биологические системы характеризуются большей или меньшей способностью к саморегуляции, т. е. к гомеостазу.
Саморегуляция – необходимое приспособление организмов для поддержания жизни в постоянно меняющихся условиях. В эволюционном развитии организмов это касается не отдельных особей, а совокупности популяции.
Популяции – открытые системы, имеющие множество каналов, по которым в популяцию поступает информация, реализуется и адекватно отвечает структурными изменениями.
Природа сигналов, воспринимаемых популяцией как “приказ” к действию, весьма разнообразна, причем система сигнализации работает безотказно.
4. Динамика популяций
Динамика популяций — это процессы изменения ее основных биологических показателей (численности, биомассы, структуры) во времени в зависимости от экологических факторов. Жизнь популяции проявляется в ее динамике — одном из наиболее значимых биологических и экологических явлений.
4.1. Кривые выживания
Кривая выживания представляет собой график зависимости от времени числа выживших на данный момент особей для некоторой начальной группы новорожденных. Каждому биологическому виду свойственна своя характерная кривая выживания. При построении графика по оси абсцисс откладывают относительный возраст, а по оси ординат — абсолютное число выживших особей или их процент от исходного числа.
Все животные и растения подвержены старению, которое проявляется в снижении жизненной активности с возрастом после периода зрелости. Непосредственные причины смерти могут быть разными, но в основе их лежит уменьшение сопротивляемости организма к неблагоприятным факторам, например болезням.
Кривая 1 на рисунке 1 близка к идеальной кривой выживания для популяции, в которой старение — главный фактор, влияющий на смертность. Так, в современной развитой стране с высоким уровнем медицинского обслуживания и рациональным питанием большинство людей доживает до старости, но среднюю ожидаемую продолжительность жизни почти невозможно увеличить более чем до 75 лет. Поскольку даже в наиболее высокоразвитых странах детская смертность выше средней, она изображается начальным участком а, а наличие долгожителей — конечным участком б (кривая 1). Кроме того, на кривую выживания влияет фактор «случайной гибели», причины которой с возрастом могут меняться. Например, в Англии наибольшая смертность в результате автомобильных аварий приходится на возраст 20—25 лет.

Рис. 1. Кривые выживания: 1 – человек или крупные млекопитающие; 2 гидра или чернохвостый олень; 3 – устрица или рыба.
Закономерность, аналогичная кривой 1, свойственна и однолетним растениям, например злаковым, ибо на поле все растения одного вида стареют одновременно.
Кривая 3 иллюстрирует изменения в популяции, у которой смертность относительно постоянна в течение всей жизни организмов (например, 50% за определенное время). Причиной смерти преимущественно служит случай, и особи гибнут до начала заметного старения. Кривая, подобная этой, была получена, например, для «популяции» стеклянных (легко бьющихся) стаканов в кафетерии. В природе к таким видам относятся, например, устрицы, дающие огромное потомство, а также растения, размножающиеся благодаря рассеиванию большого числа семян. Большая или меньшая смертность среди молодых особей приводит к изменению крутизны опускания начальной части кривой 3.
К промежуточному типу относят кривые выживания таких видов, для которых смертность мало меняется с возрастом. В природе существует много видов птиц, ящериц, мелких млекопитающих и других организмов, имеющих кривые выживания 2, которые отличаются от прямолинейной диагонали некоторой выпуклостью (вогнутостью) или волнообразностью.
Форма кривой выживания часто меняется при изменении плотности популяции. С увеличением плотности кривая становится более вогнутой, т. е. при увеличении численности организмов их смертность возрастает.
Существуют также внутривидовые различия в кривых выживания. Причины могут быть разные и нередко связаны с полом. Так, женщины живут дольше мужчин, хотя причины этого точно не известны
4.2. Рост популяции и кривые роста
Если при незначительной эмиграции и иммиграции рождаемость превышает смертность, то популяция будет расти. Рост популяции является непрерывным процессом, если в ней существуют все возрастные группы. Скорость роста популяции при отсутствии каких-либо экологических ограничений описывает дифференциальное уравнение:
dN/dx = rN, (1)
где N — численность особей в популяции; т — время; г — константа скорости естественного прироста.
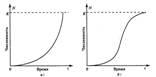 J-образная модель роста популяции. Если г 0, то со временем численность популяции становится больше. Рост происходит сначала медленно, а затем стремительно увеличивается по экспоненциальному закону, т. е. кривая роста популяции принимает J - образный вид (рис. 2, а). Такая модель основывается на допущении, что рост популяции не зависит от ее плотности. Считают, что почти любой вид теоретически способен увеличить свою численность до заселения всей Земли при достатке пищи, воды, пространства, постоянстве условий среды и отсутствии хищников. Эта идея была выдвинута еще на рубеже XVIII и XIX вв. английским экономистом Томасом Р. Мальтусом, основоположником теории мальтузианства.
J-образная модель роста популяции. Если г 0, то со временем численность популяции становится больше. Рост происходит сначала медленно, а затем стремительно увеличивается по экспоненциальному закону, т. е. кривая роста популяции принимает J - образный вид (рис. 2, а). Такая модель основывается на допущении, что рост популяции не зависит от ее плотности. Считают, что почти любой вид теоретически способен увеличить свою численность до заселения всей Земли при достатке пищи, воды, пространства, постоянстве условий среды и отсутствии хищников. Эта идея была выдвинута еще на рубеже XVIII и XIX вв. английским экономистом Томасом Р. Мальтусом, основоположником теории мальтузианства.
Рис. 2. Типы кривых роста численности популяции (модели роста популяции): а – J-образная; б – S-образная; К - поддерживающая емкость среды.
S-образная модель роста популяции. Иное развитие получает ситуация при ограниченности пищевых ресурсов либо при скоплении токсичных продуктов (отходов) метаболизма. Первоначальный экспоненциальный рост в исходных благоприятных условиях со временем продолжаться не может и постепенно замедляется. Плотность популяции регулирует истощение пищевых ресурсов, накопление токсикантов и поэтому влияет на рост численности. С увеличением плотности скорость роста популяции постепенно снижается до нуля, и кривая выходит на некоторый стабильный уровень (график образует плато). Кривая такого роста (рис. 2, б) имеет S-образную форму, и поэтому соответствующая модель развития событий называется S-образной. Она характерна, например, для дрожжей, фактором, ограничивающим их рост, является накопление спирта, а также для водорослей, самозатеняющих друг друга. В обоих случаях численность популяции не достигает уровня, на котором начинает сказываться нехватка элементов питания (биогенов).
На рост численности, в которой значительную (возможно, даже главную) роль играет пространство, также влияет перенаселенность. Лабораторные опыты с крысами показали, что по достижении определенной плотности популяции плодовитость животных резко снижается даже при избытке пищи. Возникают гормональные сдвиги, влияющие на половое поведение; чаще встречается бесплодие, поедание детенышей родителями и т. п. Резко ослабевает родительская забота о потомстве, детеныши раньше покидают гнездо, в результате чего снижается вероятность их выживания. Усиливается агрессивность животных. Подобные явления встречаются также в популяциях ряда млекопитающих, причем не только в лабораторных, но и природных условиях.
Скорость роста численности в S-образной модели определяет дифференциальное уравнение:
d/V/dx = rN(l - N/K), (2)
где К — поддерживающая емкость среды, т. е. максимальный размер популяции, которая может существовать в данных условиях, удовлетворяя свои потребности неопределенно долго.
Если N К, скорость роста отрицательна. Если N
В специализированной литературе J - и S-образные модели роста численности часто называют соответственно экспоненциальной и логистической.
Поддерживающая емкость играет решающую роль не только при росте популяции по S-образной, но также и по J-образной модели, ибо в некоторый момент времени все же наступает исчерпание какого-либо ресурса среды, т. е. он (или даже несколько одновременно) становится лимитирующим. Развитие дальнейших событий показано на рис. 4.6, а, б. После бума с внезапным выходом J-образной кривой за пределы уровня К происходит крах популяции1, т. е. катастрофа, приводящая к резкому снижению численности. Причиной краха часто бывает внезапное резкое изменение условий окружающей среды (экологических факторов), понижающее поддерживающую емкость среды. Тогда огромное число особей, не способных эмигрировать, погибает.
При наиболее благоприятном для популяции стечении обстоятельств новый уровень численности соответствует поддерживающей емкости среды или, иначе говоря, кривая роста превращается из J-образной в S-образную. Однако исчерпание пищевых ресурсов может привести также к появлению и других трудностей для популяции, например к развитию болезней. Тогда численность снижается до уровня значительно более низкого, чем поддерживающая емкость среды, а в пределе популяция может даже быть обречена на вымирание.
Для S-образной модели в случаях отставания действия регулирующих механизмов по каким-либо причинам, например, в связи с затратами времени на воспроизводство или по иным причинам временное запаздывание учитывает дифференциальное уравнение:
N/dx = rNK - rN2(x - Т)/К, (3)
где Т — время, необходимое системе для реакции на внешнее воздействие.
Вычитаемое в правой части уравнения, содержащее N2, позволяет предсказать момент выхода системы из состояния равновесия в случаях, когда время запаздывания относительно велико по сравнению с временем релаксации (1/r) системы. В итоге при увеличении в системе времени запаздывания вместо асимптотического приближения к состоянию равновесия происходит колебание численности организмов относительно теоретической S-образной кривой. В случаях, когда пищевые ресурсы ограничены, популяция не достигает устойчивого равновесия, ибо численность одного поколения зависит от численности другого, что отражается на скорости репродукции и приводит к хищничеству и каннибализму. Колебания численности популяции, для которой характерны большие значения г, малое время воспроизводства х и несложный регулирующий механизм, могут быть весьма значительными.
Описанные модели роста популяции и дифференциальные уравнения предполагают, что все организмы сходны между собой, имеют равную вероятность погибнуть и равную способ способность к размножению, так что скорость роста популяции в экспоненциальной фазе зависит только от ее численности и не ограниченали роста популяции преимущественно можно наблюдать в случаях, когда тех или иных животных вселяют или они сами распространяются в новые для них районы. Тем не менее теоретические модели роста позволяют лучше понять процессы, происходящие в естественных условиях. Большинство принципов, используемых для моделирования популяций животных, применимо также и для моделирования популяций растений.
Следует отметить, что при любой модели (как J-, так и S-образной) вначале характерна фаза экспоненциального роста численности популяции. Поэтому при сочетании благоприятных (оптимальных) значений всех факторов среды возникает «популяционный взрыв», т. е. особо быстрый рост популяции того или иного вида.
Миграция или расселение, так же как и внезапное снижение скорости размножения, могут способствовать уменьшению численности популяции. Расселение может быть связано с определенной стадией жизненного цикла, например с образованием семян.
Применительно к условиям реальной природной среды принято использовать понятия биотический потенциал — совокупность всех экологических факторов, способствующих увеличению численности популяции, или видовая способность к размножению при отсутствии ограничений со стороны среды, а также сопротивление среды — сочетание факторов, ограничивающих рост (лимитирующих факторов).
Любые изменения популяции есть результат нарушения равновесия между ее биотическим потенциалом и сопротивлением окружающей среды.
4.3. Колебания численности популяции
По достижении заключительной фазы роста размеры популяции продолжают колебаться от поколения к поколению вокруг некоторой более или менее постоянной величины. При этом численность одних видов изменяется нерегулярно с большой амплитудой колебаний (насекомые-вредители, сорняки), колебания численности других (например, мелких млекопитающих) имеют относительно постоянный период, а в популяциях третьих видов численность колеблется от года к году незначительно (долгоживущие крупные позвоночные и древесные растения).
В природе в основном встречаются три вида кривых изменения численности популяции: относительно стабильный, скачкообразный и циклический.
Виды, у которых численность из года в год находится на уровне поддерживающей емкости среды, имеют достаточно стабильные популяции. Такое постоянство характерно для многих видов дикой природы и встречается, например, в нетронутых тропических влажных лесах, где среднегодовое количество осадков и температура изменяются день ото дня и из года в год крайне мало.
У других видов колебания численности популяций носят правильный циклический характер. Хорошо знакомы примеры сезонных колебаний численности. Тучи комаров; поля, заросшие цветами; леса, полные птиц, — все это характерно для теплого времени года в средней полосе и сходит практически на нет зимой.
Широко известен пример циклических колебаний численности леммингов (северных травоядных мышевидных грызунов) в Северной Америке и Скандинавии. Раз в четыре года плотность их популяций становится столь высокой, что они начинают мигрировать со своих перенаселенных местообитаний; при этом массово гибнут в фиордах и тонут в реках, что до настоящего времени не имеет достаточного объяснения. Еще с глубокой древности известны циклические нашествия странствующей африканской саранчи на Евразию.
Ряд видов, таких, как енот, в основном имеют достаточно стабильные популяции, однако время от времени их численность резко возрастает (подскакивает) до наивысшего значения, а затем резко падает до некоторого низкого, но относительно стабильного уровня. Эти виды относят к популяциям со скачкообразным ростом численности.
Внезапное увеличение численности происходит при временном повышении емкости среды для данной популяции и может быть связано с улучшением климатических условий (факторов) и питания или резким уменьшением численности хищников (включая охотников). После превышения новой, более высокой емкости среды в популяции возрастает смертность и ее размеры резко сокращаются.
На протяжении истории в разных странах не раз наблюдались случаи краха популяций человека, например, в Ирландии в 1845 г., когда в результате заражения грибком погиб весь урожай картофеля. Поскольку рацион питания ирландцев сильно зависел от картофеля, к 1900 г. половина восьмимиллионного населения Ирландии умерла от голода или эмигрировала в другие страны.
Тем не менее численность человечества на Земле в целом и во многих регионах в частности продолжает расти. Люди путем технологических, социальных и культурных перемен неоднократно увеличивали для себя поддерживающую емкость планеты (рис. 4.8). По сути, они смогли изменить свою экологическую нишу за счет увеличения производства продуктов питания, борьбы с болезнями и использования больших количеств энергетических и материальных ресурсов, чтобы сделать обычно непригодные для жизни районы Земли обитаемыми.
В правой части рис. 4.8 приведены возможные сценарии дальнейшего изменения фактической численности людей на планете в случае превышения поддерживающей емкости биосферы.













Вебинар для учителей

Свидетельство об участии БЕСПЛАТНО!
Полезное для учителя

Реализация образовательных программ осуществляется с применением исключительно электронного обучения и ДОТ
