
Российская Федерация, Казань


СДЕЛАЙТЕ СВОИ УРОКИ ЕЩЁ ЭФФЕКТИВНЕЕ, А ЖИЗНЬ СВОБОДНЕЕ
Благодаря готовым учебным материалам для работы в классе и дистанционно
Скидки до 50 % на комплекты
только до
Готовые ключевые этапы урока всегда будут у вас под рукой
Организационный момент
Проверка знаний
Объяснение материала
Закрепление изученного
Итоги урока

Была в сети 24.04.2024 20:47

Ефимова Лилия Владимировна
Учитель химии и биологии
68 лет
Местоположение
Специализация
Клетка. Вопросы и ответы
Категория:
Биология
27.03.2024 19:09

Просмотр содержимого документа
«Клетка. Вопросы и ответы»

Строение клетки
Посмотри и определи!
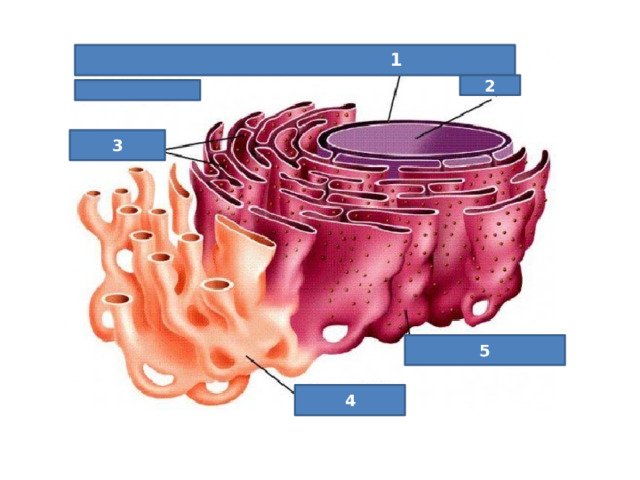
1
2
3
5
4

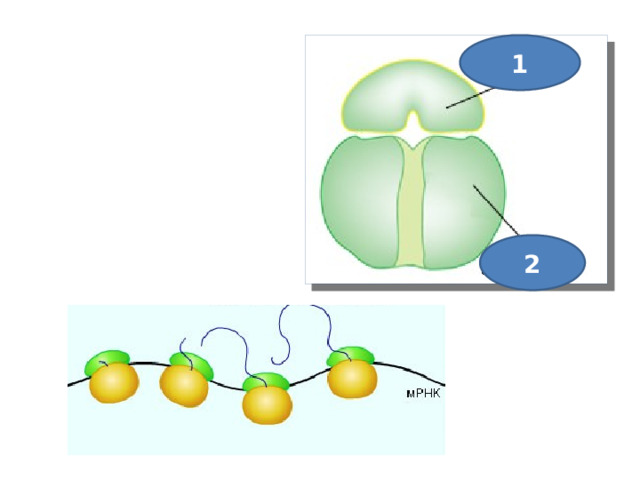
1
2

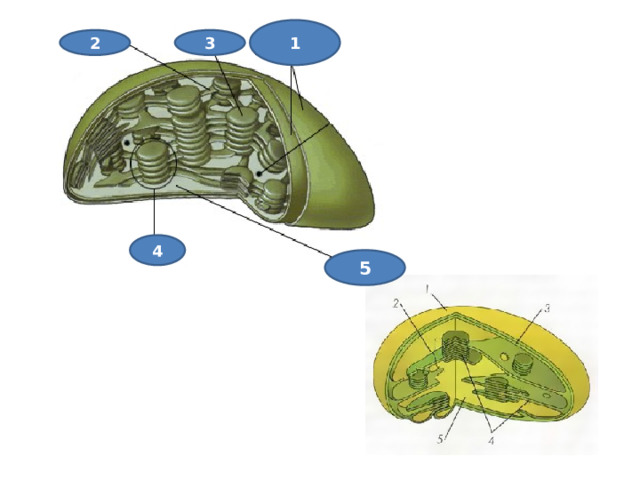
1
2
3
4
5









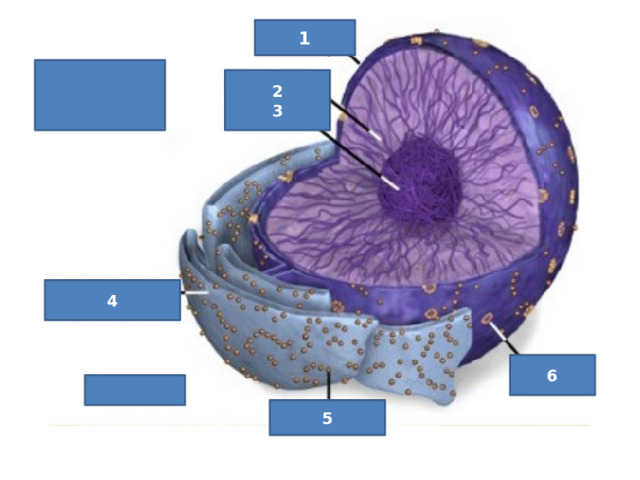
1
2
3
4
6
5
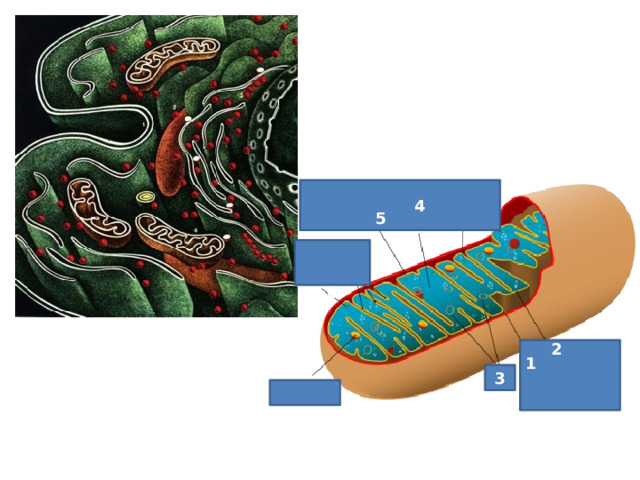
4
5
6
2
1
3

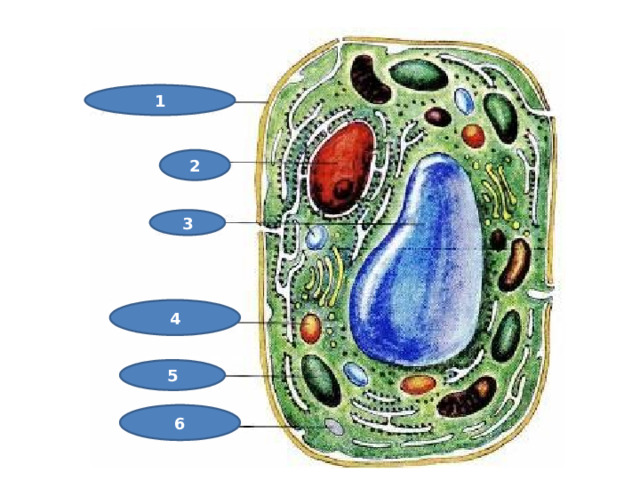
1
2
3
4
5
6

Агранулярная (гладкая) эндоплазматическая сеть( reticulum endoplasmaticum nongranulosum )
- Также представлена мембранами, образующими мелкие вакуоли и трубки, канальцы, которые могут ветвиться, сливаться друг с другом. В отличие от гранулярной эндоплазматической сети на мембранах гладкой эндоплазматической сети нет рибосом. Диаметр вакуолей и канальцев гладкой эндоплазматической сети обычно около 50—100 нм.
- Г ладкая эндоплазматическая сеть возникает и развивается за счет гранулярной эндоплазматической сети (при освобождении ее от рибосом).
- Деятельность гладкой эндоплазматической сети связана с метаболизмом липидов и некоторых внутриклеточных полисахаридов. Гладкая эндоплазматическая сеть участвует в заключительных этапах синтеза липидов. Она сильно развита в клетках, секретирующих такие категории липидов, как стероиды, например, в клетках коркового вещества надпочечников, в сустентоцитах семенников.
- Тесная топографическая связь гладкой эндоплазматической сети с отложениями гликогена (запасной внутриклеточный полисахарид животных) в гиалоплазме различных клеток (клетки печени, мышечные волокна) указывает на ее возможное участие в метаболизме углеводов.
- В поперечнополосатых мышечных волокнах гладкая эндоплазматическая сеть способна депонировать ионы кальция, необходимые для функции мышечной ткани .
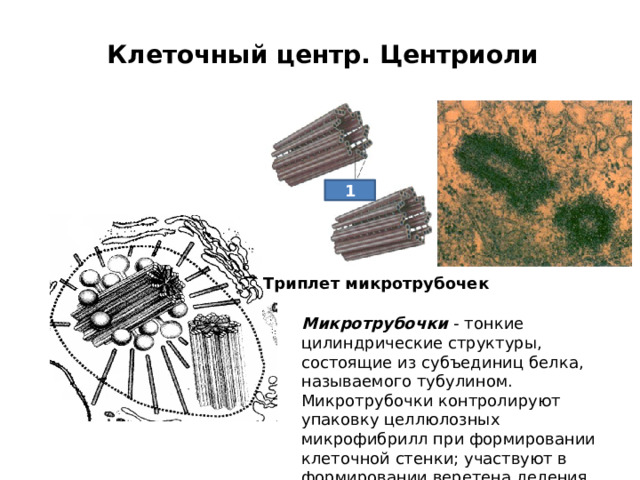
Клеточный центр. Центриоли
1
Триплет микротрубочек
Микротрубочки - тонкие цилиндрические структуры, состоящие из субъединиц белка, называемого тубулином. Микротрубочки контролируют упаковку целлюлозных микрофибрилл при формировании клеточной стенки; участвуют в формировании веретена деления.

Микрофиламенты - это длинные нити, состоящие из сократительного белка актина. Пучки микрофиламентов играют определяющую роль в токах цитоплазмы. Микрофиламенты вместе с микротрубочками образуют гибкую сеть, называемую цитоскелетом.
Цитоскелет клетки. Микрофиламенты окрашены в синий, микротрубочки – в зеленый, промежуточные волокна – в красный цвет
ЭПС (ЭПР)
Гранулярная эндоплазматическая сеть ( reticulum endoplasmaticum granulosum ) представлена замкнутыми мембранами, которые образуют уплощенные мешки, цистерны, трубочки. Ширина полостей цистерн значительно варьирует в зависимости от функциональной активности клетки. Наименьшая ширина их — около 20 нм, но они могут достигать диаметра в несколько микрометров. Отличительной чертой мембран гранулярной ЭПС является то, что они со стороны гиалоплазмы покрыты рибосомами
Рис. Строение гранулярной эндоплазматической сети. А — схема; Б — электронная микрофотография участка среза печеночной клетки. 1 — рибосомы; 2 — пластинки; 3 — внутренние полости цистерн; 4 — отщепляющиеся мембранные пузырьки, лишенные рибосом.
Рибосомы , связанные с мембранами эндоплазматической сети, участвуют в синтезе белков, выводимых из данной клетки («экспортируемые» белки). Кроме того, гранулярная эндоплазматическая сеть принимает участие в синтезе белков — ферментов, необходимых для организации внутриклеточного метаболизма, а также используемых для внутриклеточного пищеварения.
Итак, роль гранулярной эндоплазматической сети заключается в синтезе на ее рибосомах экспортируемых белков, в их изоляции от содержимого гиалоплазмы внутри мембранных полостей, в транспорте этих белков в другие участки клетки, в химической модификации таких белков и в их локальной конденсации, а также в синтезе структурных компонентов клеточных мембран.
- При рассмотрении в электронном микроскопе КГ представлен мембранными структурами, собранными вместе в небольшой зоне Отдельная зона скопления этих мембран называется диктиосомой .
- В диктиосоме КГ плотно друг к другу расположены 5—10 плоских цистерн, между которыми располагаются тонкие прослойки гиалоплазмы. Кроме них, наблюдается множество мелких пузырьков
- КГ участвует в сегрегации и накоплении продуктов, синтезированных в ЭПС, в их химических перестройках, созревании; в цистернах КГ происходит синтез полисахаридов, их связывание с белками,.
- С помощью элементов АГ происходит процесс выведения готовых секретов за пределы клетки.
- КГ обеспечивает формирование клеточных лизосом.
- Секреторная функция КГ заключается в том, что синтезированный на рибосомах экспортируемый белок отделяется и накапливается внутри цистерн ЭПС, по которым он транспортируется к зоне мембран пластинчатого комплекса. Затем накопленный белок может конденсироваться, образуя секреторные белковые гранулы (как это наблюдается в поджелудочной, молочной и других железах), или же оставаться в растворенном виде (как иммуноглобулины в плазматических клетках). В дальнейшем от ампулярных расширений цистерн КГ отщепляются пузырьки, содержащие эти белки. Такие везикулы также могут сливаться друг с другом и увеличиваться в размерах, образуя секреторные гранулы. После этого секреторные гранулы начинают двигаться к поверхности клетки, соприкасаются с плазмолеммой, с которой сливаются их собственные мембраны, и таким образом содержимое гранул оказывается за пределами клетки.
- В КГ Гольджи могут происходить многие метаболические процессы. Здесь большинство белков подвергается модификации, некоторые их аминокислоты фосфорилируются, ацетилируются или глюкозилируются.
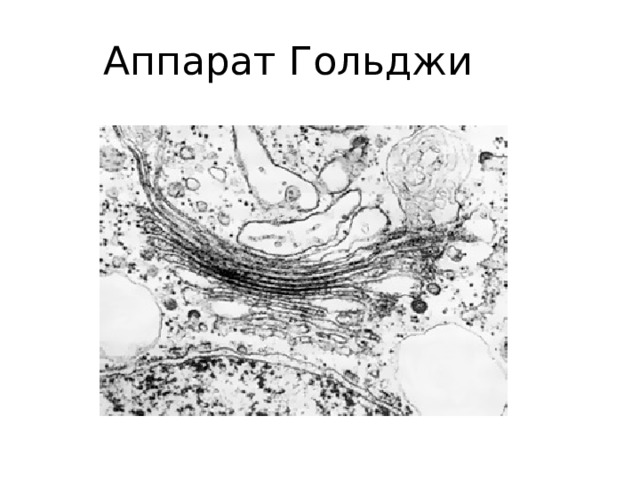
Комплекс Гольджи (внутренний сетчатый аппарат)
А — нервная клетка спинного мозга: 1 — ядро; 2 — ядрышко; 3 — комплекс Гольджи. Б — схема ультрамикроскопического строения (трехмерная реконструкция). В — комплекс Гольджи на ультратонком срезе (печеночная клетка). 1 — пузырьки; 2 — трубочки; 3 — уплощенные мешочки (цистерны); 4 — пластинки гранулярной ЭПС.
Мембраны комплекса Гольджи образуются при участии гранулярной эндоплазматической сети.
Лизосомы
- Лизосомы ( lysosomae ) — это разнообразный класс шаровидных структур размером 0,2—0,4 мкм, ограниченных одиночной мембраной. Характерным признаком лизосом является наличие в них гидролитических ферментов — гидролаз, расщепляющих различные биополимеры. Лизосомы были открыты в 1949 г. де Дювом.
- Среди лизосом можно 3 типа: первичные лизосомы, вторичные лизосомы (фаголизосомы и аутофагосомы) и остаточные тельца.
- Первичные лизосомы - мелкие мембранные пузырьки размером около 0,2—0,5 мкм, заполненные бесструктурным веществом, содержащим гидролазы.
- Вторичные лизосомы , или внутриклеточные пищеварительные вакуоли, формируются при слиянии первичных лизосом с фагоцитарными вакуолями (фагосомами) или пиноцитозными вакуолями, образуя фаголизосомы, или гетерофагосомы.
- Первичные лизосомы могут сливаться с измененными органеллами самой клетки, подвергающимися перевариванию (т.н. аутофагосомы).
- Однако расщепление, переваривание биогенных макромолекул внутри лизосом может идти в ряде клеток не до конца. В этом случае в полостях лизосом накапливаются непереваренные продукты. Такая лизосома носит название «телолизосома», или остаточное тельце ( corpusculum residuale ).
- Строение лизосом. А — схема участия структур клетки в образовании лизосом и во внутриклеточном пищеварении:
- 1 — образование из гранулярной эндоплазматической сети мелких пузырьков, содержащих гидролитические ферменты;
- 2— перенос ферментов в комплекс Гольджи;
- 3 — образование первичных лизосом;
- 4 — выделение и использование (5) гидролаз при внеклеточном расщеплении;
- 6 — эндоцитозные пузырьки;
- 7 — слияние первичных лизосом и эндоцитозных пузырьков;
- 8 — образование вторичных лизосом (фаголизосом);
- 9 — телолизосомы;
- 10 — экскреция остаточных телец;
- 11 — слияние первичных лизосом с разрушающимися структурами клетки;
- 12 — аутофагосома.
- Б — электронная микрофотография среза вторичных лизосом (обозначены стрелками).

Митохондрии
- Митохондрии— органеллы синтеза АТФ . Их основная функция связана с окислением органических соединений и использованием освобождающейся при распаде этих соединений энергии для синтеза молекул АТФ. Митохондрии часто называют энергетическими станциями клетки, или органеллами клеточного дыхания.
- Термин «митохондрия» был введен Бенда в 1897 г. для обозначения зернистых и нитчатых структур в цитоплазме разных клеток. Митохондрии можно наблюдать в живых клетках, они могут перемещаться, сливаться друг с другом, делиться. Форма и размеры митохондрий животных клеток разнообразны, но в среднем толщина их около 0,5 мкм, а длина — от 1 до 10 мкм. Подсчеты показывают, что количество их в клетках сильно варьирует — от единичных элементов до сотен. Так, в клетке печени они составляют более 20% общего объема цитоплазмы и содержат около 30—35% общего количества белка в клетке. Площадь поверхности всех митохондрий печеночной клетки в 4—5 раз больше поверхности ее плазматической мембраны.
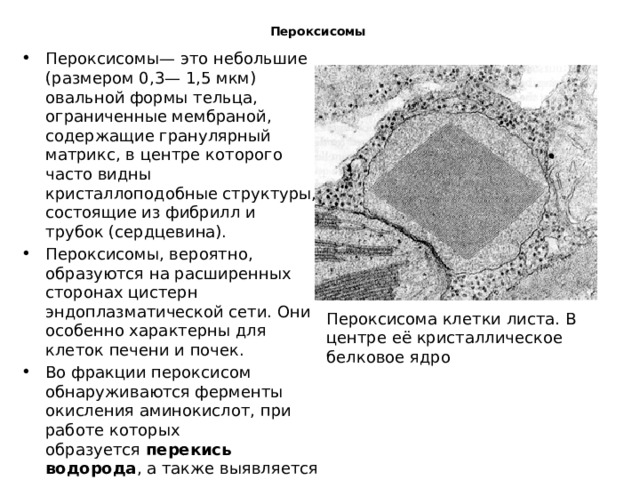
Пероксисомы
- Пероксисомы— это небольшие (размером 0,3— 1,5 мкм) овальной формы тельца, ограниченные мембраной, содержащие гранулярный матрикс, в центре которого часто видны кристаллоподобные структуры, состоящие из фибрилл и трубок (сердцевина).
- Пероксисомы, вероятно, образуются на расширенных сторонах цистерн эндоплазматической сети. Они особенно характерны для клеток печени и почек.
- Во фракции пероксисом обнаруживаются ферменты окисления аминокислот, при работе которых образуется перекись водорода , а также выявляется фермент каталаза , разрушающая ее.
- Каталаза пероксисом играет важную защитную роль, так как Н2O2 является токсическим веществом для самой клетки.
Пероксисома клетки листа. В центре её кристаллическое белковое ядро
Аппарат Гольджи
Включения
Клеточными включениями являются запасные и экскреторные вещества.
К запасным веществам относят запасные белки, жиры и углеводы. Эти вещества накапливаются в течение вегетационного периода в семенах, плодах, подземных органах растения и в сердцевине стебля.
Запасные белки, относящиеся к простым белкам - протеинам, чаще откладываются в семенах. Осаждающиеся белки в вакуолях образуют зерна округлой или эллиптической формы, называемые алейроновыми.
Запасные липиды обычно располагаются в гиалоплазме в виде капель и встречаются почти во всех растительных клетках. Это основной тип запасных питательных веществ большинства растений: наиболее богаты ими семена и плоды.
Углеводы входят в состав каждой клетки в виде растворимых в воде сахаров (глюкозы, фруктозы, сахарозы) и нерастворимых в воде полисахаридов (целлюлозы, крахмала). Главнейшим и наиболее распространенным является крахмал. Он образуется в хлоропластах. Ночью, при прекращении фотосинтеза, крахмал гидролизуется до сахаров и транспортируется в запасающие ткани - клубни, луковицы, корневища. Там в особых типах лейкопластов - амилопластах - часть сахаров откладывается в виде зерен вторичного крахмала.


Экскреторные вещества (продукты вторичного обмена)
- К клеточным включениям относятся и экскреторные вещества, например кристаллы оксалата кальция (одиночные кристаллы, рафиды - игольчатые кристаллы, друзы - сростки кристаллов, кристаллический песок - скопление множества мелких кристаллов). Реже кристаллы состоят из карбоната кальция или кремнезема (цистолиты ).
- В отличие от животных, выводящих избыток солей вместе с мочой, растения не имеют развитых органов выделения. Поэтому считается, что кристаллы оксалата кальция являются конечным продуктом метаболизма протопласта, образующимся как приспособление для выведения из обмена излишков кальция. Как правило, эти кристаллы накапливаются в органах, которые растение периодически сбрасывает (листья, кора).
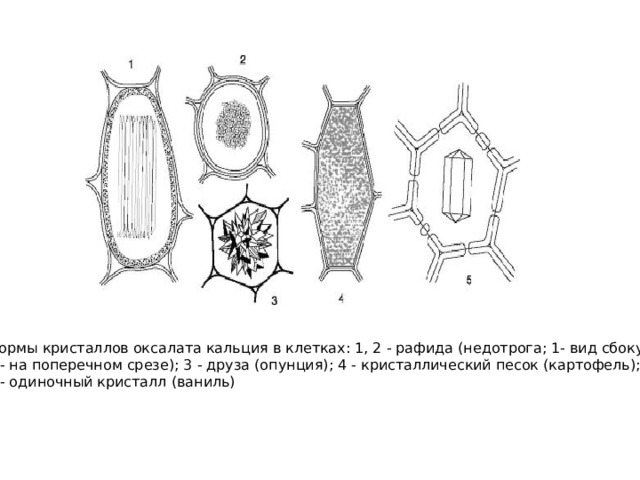
Формы кристаллов оксалата кальция в клетках: 1, 2 - рафида (недотрога; 1- вид сбоку,
2 - на поперечном срезе); 3 - друза (опунция); 4 - кристаллический песок (картофель);
5 - одиночный кристалл (ваниль)

Клеточная стенка
- Целлюлозный каркас клеточной стенки заполнен нецеллюлозными молекулами матрикса. В состав матрикса входят полисахариды, называемые гемицеллюлозами; пектиновые вещества (пектин), очень близкие к гемицеллюлозам, и гликопротеиды. Пектиновые вещества, сливаясь между соседними клетками, образуют срединную пластинку, которая располагается между первичными оболочками соседних клеток.
- При растворении или разрушении срединной пластинки (что происходит в мякоти созревших плодов) возникает мацерация (от лат. maceratio - размягчение). Естественную мацерацию можно наблюдать у многих перезрелых плодов (арбуз, дыня, персик). Искусственную мацерацию (при обработке тканей щелочью или кислотой) используют для приготовления различных анатомических и гистологических препаратов.
- Клеточная стенка в процессе жизнедеятельности может подвергаться различным видоизменениям - одревеснению, опробковению, ослизнению, кутинизации, минерализации

Характерные изменения вторичной клеточной стенки

- Какие органоиды или части клетки не были названы?













Вебинар для учителей

Свидетельство об участии БЕСПЛАТНО!
Полезное для учителя

Реализация образовательных программ осуществляется с применением исключительно электронного обучения и ДОТ


